Категории:
ДомЗдоровьеЗоологияИнформатикаИскусствоИскусствоКомпьютерыКулинарияМаркетингМатематикаМедицинаМенеджментОбразованиеПедагогикаПитомцыПрограммированиеПроизводствоПромышленностьПсихологияРазноеРелигияСоциологияСпортСтатистикаТранспортФизикаФилософияФинансыХимияХоббиЭкологияЭкономикаЭлектроника
Роль ДНК в сохранении, передачи и реализации наследственной информации.
Передача генетической информации в клетке основана на матричных процессах (репликации, транскрипции, трансляции). Синтез дочерней цепи (репликация) молекулы ДНК происходит по матрице одной из двух родительских цепей с образованием новой молекулы ДНК. Синтез молекулы РНК совершается в процессе транскрипции ДНК по матрице одной из двух цепей ДНК. Такая матричная (информационная) РНК может рассматриваться как посредник между ДНК и белком. Далее при синтезе белков генетическая информация, закодированная в последовательности триплетов азотистых оснований (канонов), транслируется в аминокислотную последовательность полипептидных цепей. Во время репликациипроисходит расхождение двух цепей ДНК, и каждая из них служит матрицей для синтеза дочерней цепи. Такой способ репликации называется полуконсервативным. При этом дезоксирибонуклеотиды встраиваются в дочернюю цепь согласно правилу комплементарности азотистых оснований (А — Т, G — С). Вновь образованная молекула состоит из одной родительской и одной дочерней цепи ДНК. Образование дочерних хромосом происходит на стадии синтеза (S) в интерфазе между митотическими делениями и перед первым делением мейоза, В анафазе удвоенные хромосомы расходятся по дочерним клеткам. Таким образом, без процесса репликации невозможно сохранение диплоидного числа хромосом в соматических клетках и образование гаплоидного набора хромосом в половых клетках после двух делений мейоза. Однако при делении клеток происходит не только сохранение числа хромосом, но и воспроизведение последовательности азотистых оснований в молекулах ДНК, основанное на комплементарности пар оснований родительской и дочерней цепей ДНК.Репарация. Система зашиты клетки включает различные типы репарации поврежденной молекулы ДНК. Этот процесс может быть одноэтапным и многоэтапным, происходить как на свету, так и в темноте. Ошибки репликации исправляют специальные ферменты с корректирующей функцией; они находят и удаляют некомплементарное основание. Затем происходит замена на основание, соответствующее правилу комплементарноста (А- Т, G - С). Рекомбинация.Образование новых сочетании генов происходит и результате обмена участками между гомологичными последовательностями ДНК (кроссинговер). В процессе кроссинговера происходит обмен участками между гомологичными хромосомами. При этом, чем дальше расположены гены, тем более часто происходит между ними кроссинговер. Этот принцип был использован при построении первых генетических карту дрозофилы и кукурузы. Рекомбинация может иметь место и при незначительной гомологии нуклеотидных пар, например, при интеграции фагов в хромосому бактерий, и при фактическом отсутствии этой гомологии в случае перемещения мобильных диспергированных элементов по эукариотическим хромосомам. Последние два типа событий относят к незаконной рекомбинации. Транскрипция. Генетическая информация, записанная в последовательности оснований в молекуле ДНК, передается на молекулу рибонуклеиновой кислоты (РНК) в процессе транскрипции. РНК отличается от ДНК наличием в сахарофосфатном остове молекулы сахара рибозы вместо дезоксирибозы и другого азотистого основания - урацила (вместо тимина), комплементарного аденину. Транскрипция ДНК -матричный процесс, во время которого молекула РНК синтезируется по матрице одной из двух цепей ДНК. При этом происходит локальное расплетение цепей ДНК в транскрибируемом участке и присоединение рибонуклеотидных остатков к растущей цепи РНК. По окончании транскрипции каждого очередного участка молекулы ДНК ее двухспиральная структура восстанавливается. Транскрипция заканчивается на терминаторных последовательностях гена с отделением сиинтезированной одноцепочечной молекулы РНК. В процессе транскрипции участвуют не только специальные ферменты, но и многочисленные регуляторные белки. Такие белки взаимодействуют с регуляторными последовательностями генов, обеспечивая процесс начала и окончания транскрипции и уровень нарабатываемого первичного продукта. Трансляция. Передача генетической информации с мРНК на белок носит название трансляции. Биосинтез белка происходит на цитоплазматических структурах, называемых рибосомами. Рибосома, продвигается вдоль мРНК, последовательно выбирая из среды те аминокислоты, соединенные с транспортными РНК (тРНК), которые соответствуют кодирующим последовательностям нуклеотидов. При этом последовательность кодонов в зрелой молекуле РНК определяет последовательность аминокислот в полипептидной цепи.
45.Строение ДНК И РНК. Видовая специфичность ДНК.
ДНК-стойкое хим. соединение, не подвергающейся воздействию спиртов, щелочей, кислоты. Днк можно разрезать на часть и эти части могут склеиваться и соед. между собой. ДНК способна денотурации – т.е. раскручиванию спирали под влиянием ферментов – происходит при репликации и транскрипции. После прекращения воздействия ферментов, ДНК способна восстанавливать первичную и вторичную стр-ру - ренотерация (восстановление). ДНК находится в ядре клетки.
ДНК состоит из 2 цепочек закрученных в спираль(внутри азотистое основание, сахар, фосфорные остатки.)
ДНК является первичным носителем наследственной информации. Это означает, что вся информация о структуре, функционировании и развитии отдельных клеток и целостного организма записана в виде нуклеотидных последовательностей ДНК.
РНК относится к нуклииновым к-ам, является полимером. РНК состоит из 1 цепочки и содержит сахар – рибозу. Состав каждой нуклеотида входит: Сахар – рибоза, фосфорные к-ты,  - азотистого основания(Аденин, гуанин, цитозин и уроцил.). РНК меньше чем ДНК по молек. массе и поэтому может существовать в разных формах, подвергаться действию кислот, щелочей, ферментов.
- азотистого основания(Аденин, гуанин, цитозин и уроцил.). РНК меньше чем ДНК по молек. массе и поэтому может существовать в разных формах, подвергаться действию кислот, щелочей, ферментов.
Существует несколько типов РНК:
1 Информационная (Матричная)-в виде цепочки
2 Рибосомная
3.Транспортная(центральная петля имеет 3 нуклеотида, в виде трехлистника)
Матричная РНК представлена в виде цепочки- в первичной стр-ре. Составляет 5% клеточной РНК. Служит для передачи генетической информации от ДНК на рибосомы при биосинтезе белка. Для защиты матр. в клетке образ. столько типов матр. РНК сколько типов белков существует в клетки. Иногда на рибосомах матр. РНК может присоединиться нескольким рибосомам и образовать несколько белков – назыв. полисомы.
Для защиты матр. РНК от воздействия ферментов на передних 5 штрихах в конце – колпачок. На 3 штрихе в конце находится – аденин нуклеотид.
Транспортная РНК – Составляет 10% клеточной РНК. Включает до 80 нуклеотидов, некоторые участки компелементируются друг с другом. Центральная петля имеет 3 нуклеотида, которые комплементирует нуклеотид на матричной РНК. В клетке находится около 60 видов транспортного РНК. Функция- Доставка аминокислот к рибосомам. Транспортная РНК синтезируется в ядре клетки, к ней присоединяется белок- называется EF-фактор.
Рибосомная РНК- ее больше всего в клетке, около 80%, входит в состав рибосом. По молекулярной массе могут быть: 1. Большие молекулы РНК – от 1600 до 4700 нуклеотидов. 2. Малые – от 120 до 160. Образуется рибосомная РНК в области вторичных перетяжек хромосом, которые в интерфазу образует ядрышек. Синтез происходит в ядрышках.
46. Пространственная модель строение молекулы ДНК ,предложенная Уотсоном и Криком.
1953г- Уотсон и Крик использовали закономерность и ренгеноструктивный анализ, который сделала Р.Франклинк ,они построили модель молекулы ДНК.
1.ДНК состоит из 2 цепочек закрученных в спираль(внутри азотистое основание, сахар, фосфорные остатки.)
2.Азотистые основание повернуты внутрь спирали и удерживаются в таком состоянии за счет водородных связей между азот. оснований.
3 Азот. основания коплементарны друг другу.
4. Каждая цепочка ДНК включает 100000 нуклиотидов. Диаметр спирали = 20 акстрен.
5. Один виток спирали встречает 10 пар азот. основ. Или 10 пар нуклеотидов. Длина витка = 34 акстренам.
6. Цепочки антипаралелльны друг к другу и имеют противоположное направленность.
7. Азот. основ. расположены внутри спирали под прямым углом к оси. Сахара находятся под прямым углом к азотистым основаниям.
8. На последующих азот. основ. не накладывает никаких ограничений.
ДНК имеет первичную стру-ру – это последовательность нуклеотидов. Образующийся спирали – вторичная стру-ра. ДНК образует суперветки – это третичная стру-ра.
47. Схема репликации(самоудвоения) ДНК.
Процесс репликации основан на 2-ух принцыпах:
1. Новая молекула ДНК не создается заново, а синтетически происходит на основе матрицы ,для каждого новая молекула ДНК является каждой цепочкой исходного молекулы ДНК.
2. Такой синтез возможен благодаря принципу комплементарности. Механическая реплекация ДНК предложена Уотсоном и Криком.
Происходит раскручивании спирали ДНК. Под ферментом топоизомераза цепочки ДНК отходят друг от друга ,и вместе расходятся цепочеки образуя репликационную вилку. Синтез идет антипараллельно. 1 цепочка – Под начало ДНК к реплекационной вилке, 2цепочка – синтез идет медленно, идет от вилке к началу и синтезируется фрагментами. Фрагменты включают до 1000 нуклеотидов. 2 цепь отстоющая. Схема получается полуконсирвативная – т.к. одна цепь старая, другая новая. ДНК-полимеразе, исходный субстрат для синтеза новой цепи ДНК является дезоксирибонуклеозинтрифасфаты. Образующийся новые цепочки обязаны в начале синтеза образовывать несколько нуклеотидов образуя – активатор, именно к этому куску и подсоединяется ДНК. Отстающая цепь(запоздалая) – синтезирует фрагмент.
В синтезе ДНК 3 этапа:
1 Инициация – Трансляция происходит на рибосоме. И-РНК присоединяется к микрочастицам, 1 кадон – инициатор(АУГ) без него не будет происходить синтез. Подходит транспортная РНК(строение как клевер).Антикодон на инфРНК должен соответствовать кодону, потом присоединяется микрочастица.
2 Элонгация – наращивания цепочек из нуклеотида в новые цепи по принципу комплементарности к старой цепи. Рибосома движется по и-РНК и поступает новый кодон. Доставляет аминокислоту.
3 Терминация – Последний кодон терминатор не кодирует аминокислоту, синтез прекращается.
48. Типы РНК и их функции в процессе синтеза белка.
РНК относится к нуклииновым к-ам, является полимером. Состав каждой нуклеотида входит: Сахар – рибоза, фосфорные к-ты, - азотистого основания(Аденин,гуанин,цитозин и уроцил.). РНК меньше чем ДНК по молек. Массе и по этому может существовать в разных формах, подвергается действию кислот, щелочей, ферментов.
Существует несколько типов РНК:
1 Матричная
2 Информационная ( передает в цитоплазму генетическую информацию от ДНК, находящейся в ядре)
3.Транспортная
4.Рибосомная
Матричная РНК представлена в виде цепочки- в первичной стр-ре. Для защиты матр. В клетке образ. столько типов матр. РНК сколько типов белков существует в клетки. Иногда на рибосомах матр. РНК может присоединиться нескольким рибосомам и образовать несколько белков – назыв. полисомы.
Транспортная РНК
Включает до 80 нуклеотидов, некоторые участки коплементраности друг к другу. Центральная петля имеет 3 нуклеотида которые комплементируют нуклеотид на матричном РНК.В клетки около 60 видов транспортной РНК. Ф-ция трансп. РНК: Доставка аминокислот к рибосомам. Трансп. РНК синтезируется в ядре клетки, к ней присоединяется белок – называется EF-фактор.
Рибосомная РНК
Ее больше всего в клетки, около 80%, входит в состав рибосом. У эукариот- существует 5 видов, у прокариот- 3 вида. По молекулярной массе может быть: 1 Большие молекулы РНК- от 1600 до 4700 нуклеотидов.2 Малые -от 120 до 160 нуклеотидов. Образуются рибосомные РНК в области вторичных перетяжек хромосом, которые в интерфазе образуют ядрышек. Синтез происходит в ядрышках.
49. Синтез белка в клетки. Транскрипция и трансляция.
В составе белка входят 20 аминокислот (незаменимые и заменимые). Каждая аминокислота имеет корбоксильную группу (СООН), аминогруппу( NH2) и отличаются друг от друга только радикалами. Белки молекул образуются в результате образования пептидной связи между соседними аминокислотами.
Белки имеют стр-ры:
1. Первичная- это последовательная аминокислота в молекуле ДНК.
2. Вторичная- это вид спирали, за счет образования водородных связей между аминогрупп и карбоксильных групп на соседних белках.
3.Третичная – более плотная упаковка спирали в виде глобулы, за счет дисульфидных связей между радикалами аминокислот и белки имеют 4-ую стру-ру(несколько молекул одного и того же белка объединяются для выполнения своей функции)
Белки способны к денатурации – потери 2-ой,3-ей стру-ры под влиянием температуры, освещения, обезвоживания, облучения. Если теряется 1-ая стр-ра, то белок не способен восстанавливать свои фу-ции.
Функции белка:
1 Ферментативная фу-ция;
2 Строительная – белки входят в мембрану клетки.
3 Двигательная – сократительные белки в мышечных волокнах.
4 Транспортная – гемоглобин переносит кислород.
5 Защитная
6. Энергетическая – при расщеплении белков выделяется энергия.
Трансляция(синтез белка)– это передача наследственной информации с информационной ДНК на последовательные аминокислоты в белке (синтез белка на рибосоме)
Происходит процесс на рибосоме, обязательный компонент информационной ДНК, рибосомной РНК, более 20 видов ферментов и более 50 видов белков, которые на разных этапах появляются.
Этапы трансляции:
1) Происходит активация аминокислот, за счет взаимодействия аминокислот с молекулой АТФ и происходит процесс с затратой энергии.
2) Активируется аминокислоты, присоединяется к транспортной РНК.
3) Присоединяется белок - TF3 и с помощью этого белка рибосома узнает 1-кадон на инф. РНК.
1-3 этапа- транскрипция, с 4 этапа начинается трансляция.
4) Второй этап- элонгация, происходит наращивание цепочки из аминокислот. Присоединяется 2 частицы рибосом. Инф. РНК сдвигается, а в активный центр рибосомы поступает кадон. В центре образуется цепочка из аминокислот.
5) Последний этап – терминация, т.е. на этом этапе в активном центре поступает 3 кадона темина: УАА,УАГ,УГА. Синтез белка заканчивается.
Иногда на 1 инф. РНК содятся много рибосом, образуя сразу несколько молекул одного и того же белка.
Транскрипция– это считывания или перенос насл.инф. с ДНК на инф. РНК.
Матрицей для синтеза РНК явл. 1 из цепочек ДНК – смысловая цепочка. В основе транскрипции лежит процесс копирования на основе принципа камплиментраности.
Транскрип. + Трансляц. = Экспрессия гена или реализация фун-ции гена. В клетки насл. инф. передается от ДНК,РНК и далее на белок.
50. Генетический код и его свойства. Понятие о триплете, кодоне и антикодоне.
Генетический код(код наследования)-это определенное соответствие между последование нуклеотидов ДНК и последовательных аминокислот в молекуле белка. Свойства генетического кода:
1. Генетический код триплетен - три нуклеотида в молекуле ДНК кодирует определенную аминокислоту в молекуле белка.
2. Вырожденность- проявляется всего аминокислот -20, разновидных нуклеотидов-4, сочетаний-64 из них 61-смысловые кодоны, и 3 кодон- кодоны КЕВИРНАТОРЫ (УАА, УАД, АУД)
3. Генетический код универсален - независимо от уровня организации генетический код у всех одинаковый.
4. Генетический код не перекрывающейся – триплеты считываются один за другим без промежутков.
5. Кодон АУГ является стартовым кодоном – если находится в середине, то кодирует аминокислоту – метионин.
6. В конце находится 1 из 3 кодонов терминатор или стопкодон (нонсенс)
Функция:
Воспроизводить наследственную информацию, репликация(редупликация)
Триплет (лат. triplus — тройной) — в генетике, комбинация из трёх последовательно расположенных нуклеотидов в молекуле нуклеиновой кислоты.
В информационных рибонуклеиновых кислотах (и-РНК) триплеты образуют так называемые кодоны, с помощью которых в и-РНК закодирована последовательность расположения аминокислот в белках.В молекулах транспортных РНК (т-РНК) триплеты образуют антикодоны.
Кодон (кодирующий тринуклеотид) — единица генетического кода, тройка нуклеотидных остатков (триплет) в ДНК или РНК, обычно кодирующих включение одной аминокислоты. Последовательность кодонов в гене определяет последовательность аминокислот в полипептидной цепи белка, кодируемого этим геном.
Антикодон — триплет (тринуклеотид), участок в транспортной рибонуклеиновой кислоте (т-РНК), состоящий из трёх неспаренных (имеющих свободные связи) нуклеотидов. Спариваясь с кодоном матричной РНК (м-РНК), обеспечивает правильную расстановку каждой аминокислоты при биосинтезе белка.
51. Регуляция синтеза белка в клетке (схема Жакоба и Моне)
Основным условием существования любых живых организмов является наличие тонкой, гибкой, согласованно действующей системы регуляции, в которой все элементы тесно связаны друг с другом. В белковом синтезе не только количественный и качественный состав белков, но и время синтеза имеют большое значение. От этого зависит приспособление микроорганизмов к условиям окружающей питательной среды как биологической необходимости или приспособление сложного многоклеточного организма к физиологическим потребностям при изменении внутренних и внешних условий. Клетки живых организмов обладают способностью синтезировать огромное количество разнообразных белков. Однако они никогда не синтезируют все белки. Количество и разнообразие белков, в частности ферментов, определяются степенью их участия в метаболизме. Более того, интенсивность обмена регулируется скоростью синтеза белка и параллельно контролируется аллостерическим путем. Таким образом, синтез белка регулируется внешними и внутренними факторами и условиями, которые диктуют клетке синтез такого количества белка и такого набора белков, которые необходимы для выполнения физиологических функций. Все это свидетельствует о весьма сложном, тонком и целесообразном механизме регуляции синтеза белка в клетке.
Общую теорию регуляции синтеза белка разработали французские ученые, лауреаты Нобелевской премии Ф. Жакоб и Ж. Моно. Сущность этой теории сводится к «выключению» или «включению» генов как функционирующих единиц, к возможности или невозможности проявления их способности передавать закодированную в структурных генах ДНК генетическую информацию на синтез специфических белков. Эта теория, доказанная в опытах на бактериях, получила широкое признание, хотя в эукариотических клетках механизмы регуляции синтеза белка, вероятнее всего, являются более сложными. У бактерий доказана индукция ферментов при добавлении в питательную среду субстратов этих ферментов. Добавление конечных продуктов реакции, образование которых катализируется этими же ферментами, напротив, вызывает уменьшение количества синтезируемых ферментов. Это последнее явление получило название репрессии синтеза ферментов. Оба явления – индукция и репрессия – взаимосвязаны.
Согласно теории Ф. Жакоба и Ж. Моно, в биосинтезе белка у бактерий участвуют по крайней мере 3 типа генов: структурные гены, ген-регулятор и ген-оператор.
Структурные гены определяют первичную структуру синтезируемого белка. Именно эти гены в цепи ДНК являются основой для биосинтеза м-РНК, которая затем поступает в рибосому и, как было указано, служит матрицей для биосинтеза белка. Синтез м-РНК на структурных генах молекулы ДНК непосредственно контролируется определенным участком, называемым геном-оператором. Он служит как бы пусковым механизмом для функционирования структурных генов.
Ген-оператор локализован на крайнем отрезке структурного гена или структурных генов, регулируемых им. «Считывание» генетического кода, т.е. формирование м-РНК, начинается с промотора – участка ДНК, расположенного рядом с геном-оператором и являющегося точкой инициации для синтеза м-РНК, и распространяется последовательно вдоль оператора и структурных генов. Синтезированную молекулу м-РНК, кодирующую синтез нескольких разных белков, принято называть полигенным (полицистронным) транскриптом. Координированный одним оператором одиночный ген или группа структурных генов образует оперон. В свою очередь деятельность оперона находится под контролирующим влиянием другого участка цепи ДНК, получившего название гена-регулятора.
Структурные гены и ген-регулятор расположены в разных участках цепи ДНК, поэтому связь между ними, как предполагают Ф. Жакоб и Ж. Моно, осуществляется при помощи вещества-посредника, оказавшегося белком и названного репрессором. Образование репрес-сора происходит в рибосомах ядра на матрице специфической мРНК, синтезированной на гене-регуляторе. Репрессор имеет сродство к гену-оператору и обратимо соединяется с ним в комплекс. Образование такого комплекса приводит к блокированию синтеза мРНК и, следовательно, синтеза белка, т.е. функция гена-регулятора состоит в том, чтобы через белок-репрессор прекращать (запрещать) деятельность структурных генов, синтезирующих мРНК. Репрессор, кроме того, обладает способностью строго специфически связываться с определенными низкомолекулярными веществами, называемыми индукторами, или эффекторами. Если такой индуктор соединяется с репрессором, то последний теряет способность связываться с геном-оператором, который, таким образом, выходит из-под контроля гена-регулятора, и начинается синтез мРНК. Это типичный пример отрицательной формы контроля, когда индуктор, соединяясь с белком-репрессором, вызывает изменения его третичной структуры настолько, что репрессор теряет способность связываться с геном-оператором. Процесс этот аналогичен взаимоотношениям алло-стерического центра фермента с эффектором, под влиянием которого изменяется третичная структура фермента и он теряет способность связываться со своим субстратом.
52. Современное представление о строении и функции гена. Ген-регулятор, оперон, структурные гены.
Гены - есть структурные гены, которые контролируют синтез ферментативного расщепления лактозы до глюкозы. Существуют 3 вида этих ферментов(структурных генов) – они вырабатывают ферменты, работы этих генов контролируется геном оператором. Оператор работает по принципу выключателя ( вкл. или выкл. эти гены). Перед оператором находится активатор – на нем начинается транскрипция. В конце находится ген терминатор и вся эта система называется ОПЕРОН – эта система находится под влиянием гена регулятора он выделяет белок регулятор.
Строение генов:
Ген – участок молекулы ДНК, кодирующая последовательность аминокислот в молекуле белка.
Специфичность гена – проявляется в числе и последовательных нуклеотидах. В среднем каждый ген включает около 1000 нуклеотидов.
Выделают структурные гены – кодируют синтез белка, имеют мозаичное строение, т.е. имеет активатор (экзоны) и не активаторы (интроны) активных зон.
У прокариот имеются только активные зоны. Имеются гены контролирующие синтез рРНК,тРНК и синтез регуляторов белков. Гены эукариот имеют много избыточного ДНК (не несущую информацию) и находятся такие участки в гетерохроматиновых участках хромосом. В ДНК имеются повторы (т.е. один и тот же ген может повторяться и кодировать ту же аминокислоту) Повторы регулируют транскрипцию и трансляцию и выполняют защитную функцию.
Промотор (перед оператором)включает 24 нуклеотида, на него действуют гены усилители или ослабители транскрипции. В состав ДНК входят мобильные мигрированные элементы – это фрагменты ДНК меняющие свое местоположение (прыгающие гены).
54. Понятие о норме реакции организма на условии внешней среды.
Нормой реакции генотипа – называют проявляющийся признак в конкретных условиях среды.
Норма реакции генотип показывает размах изменчивости признака в конкретных условиях среды. Определяет возможности генотипа.
Признаки нормы реакции:1) Широка, 2) Узкая.
Менее значимым для организма признака больше подвергается влиянию фактору среды, чем более значимый для организма, тем в меньшей степени зависит от среды и имеет узкую норму реакции.
Не наследственной изменчивости относят фенокопии – по фенотипу имитируют наследственные изменения.
Не наследственные изменения относят морфозы – возникает в критические периоды эмбриогенеза.
Так, у китайской примулы окраска цветков варьирует от белой (при температуре 30°С) до розовой (при 20 °С); у бабочек траурниц, развивающихся летом (при высоких температурах), белая кайма на крыльях четко очерчена, а у развивающихся весной (при пониженных температурах) она размыта. Изменения фенотипа в пределах обусловленной генотипом Н. р. могут возникать в ответ на любые колебания условий среды, в которой протекает развитие организма. Наблюдаемые изменения часто глубоко меняют фенотип, но не затрагивают генотип, т.к. они обратимы: при возвращении исходных условий среды организм либо в том же поколении (загар человека, густота шерсти млекопитающих, окраска цветков примулы), либо в следующем (окраска крыльев траурницы, число стеблей у одного растения пшеницы), а иногда и в ряду поколений (т. н. длительной модификации) возвращается к первоначальному состоянию, утрачивая признаки, возникшие при изменении условий обитания.
Другим доказательством того, что изменения в пределах Н. р. происходят без изменений генотипа, служит их наличие и в чистых линиях т. е. генотипически однородном материале. Более или менее широкая Н. р. вырабатывается в процессе естественного отбора ; она присуща всем организмам, обеспечивая их выживание при сдвигах условий обитания. Т. о., генотип определяет не жёсткую комбинацию строго детерминированных признаков фенотипа, а именно Н. р. организма при его формировании и развитии
55. Мутационная теория Г. Де-Фриза и СИ. Коржинского.
Основные положения мутационной теории Г. Де Фриза сводятся к следующему:
1. Мутации возникают внезапно как дискретные изменения признаков.
2. Новые формы устойчивы.
3. В отличие от ненаследственных изменений мутации не образуют непрерывных рядов, не группируются вокруг какого-либо среднего типа. Они представляют собой качественные изменения.
4. Мутации проявляются по-разному и. могут быть как полезными, так и вредными.
5. Вероятность обнаружения мутаций зависит от числа исследованных особей.
6. Сходные мутации могут возникать неоднократно.
56. Особенности мутационной изменчивости. Спонтанный мутагенез.
Мутационная изменчивость – наследственная изменчивость связана с изменением генотипа, т.е. с изменением генов и хромосом. 1901-1903 – Гуго и Канжинский разработали мутационную теорию изменчивости.
Мутация – это внезапное скачкообразное изменение наследственного материала.
Мутация – это качественное изменение (он или есть или его нет).
Мутация стойко передается из поколения в поколение.
Мутации не образует не прерывного рядов и не группируется вокруг среднего типа.
Мутации могут быть: Полезными, вредными, нейтральными. Одни и те же мутации могут возникать повторно. Классификация мутации: 1 Прямые – доминантные и рецессивные. 2 Обратные – рецессивные доминантные в доминирующих генах.
Мутации делят:
1) Спонтанные – самопроизвольное возникновение. Возникает редко с частотой 1*  клеток, но встречаются.
клеток, но встречаются.
2) Индуцированная – искусственная вызванная мутация. Возникает под влиянием факторов- мутагенов.
57. Понятие о точечных и хромосомных мутациях, их типы.
Гены или точковые мутации: Ген – участок молекулы ДНК. При генной мутации изменяется структура ДНК на уровне нуклеотидов.
Разновидности мутации:
1 Замена азотистого основания
транзиции 
2 Перекрестные замены
трансверсии 
3 Вставка нуклеотида
АТГЦ
Хромосомные оберации или хромосомная перестройка.
Связано эти мутации с изменением структуры хромосом.
1. Может быть потеря участка хромосом (или нехватка – дефишез)- хромосома становится короче и теряется часть генов
2. Может теряться срединный участок – дереции.
3. Удвоение определенного участка хромосома (удваиваться число генов) –дубликация, особь может быть жизнеспособной.
4. Поворот участка хромосома на 180°, при этом изменения порядка расположения генов – Инверсия, нарушается процесс половой системы.
5. Транслакация – обмен участками между не гомологичными хромосомами. Встречается робертсоновская транслокация. Особи жизнеспособны, но нарушается плодовитость.
6. Образование изохромосом– т.е. внутри хромосом может разбиваться на фрагменты и образовывать новый ген.
58. Причины, вызывающие появление мутаций. Понятие о мутагенах.
Есть много причин мутаций. Некоторые возникают спонтанно, в результате ошибок при репликации и репарации ДНК. Другие, однако, индуцируются, возникают в результате действия мутагенов или факторов окружающей среды. Вещество или воздействие называются мутагенными, если они вызывают мутации с частотами, превышающими частоты спонтанного фона.
Существует три основных типа мутагенов, или мутагенных агентов:
1) Ионизирующая радиация. Альфа, бета, гамма и рентгеновские лучи могут нарушать нормальную последовательность оснований ДНК, в основном, выбивая из нее пары оснований.
2) Неионизирующая радиация. Ультрафиолетовый свет приводит к сшивке двух расположенных рядом тиминов в нити ДНК, что блокирует репликацию ДНК, и требуется ее репарация (восстановление). Если репарация не может успешно завершиться, возникают точечные мутации.
3) Химические мутагены. Многие химические вещества взаимодействуют с ДНК таким образом, что изменяются пары оснований.
Выделяют три основных типа химических мутагенов:
· Аналоги оснований
· Модификаторы оснований
· Интеркалирующие агенты.
Большинство мутаций происходит не в половых клетках, а в соматических и называется соматическими клеточными мутациями. Если они и изменяют фенотип, то только у одного организма. Физические мутагены Химические мутагены
4) Биологические мутагены. Мутагены — химические и физические факторы, вызывающие наследственные изменения —мутации. Впервые искусственные мутации получены в1925 году Г. А. Надсеном и Г. С. Филипповым у дрожжей действием радиоактивного излучения радия
59. Классификация мутаций по генотипу и фенотипу.
Классификация мутации по фенотипу:
1) Морфологические мутации – затрагивают строение органов и тканей (карликовость)
2) Физиологические мутации – функции органов и тканей. (Изменение плодов, жизнеспособности)
3) Геохимические – нарушение синтеза определенных биоактивных веществ (нарушение обмена веществ)
Первые классификации мутации по их действию открыты в 1932 Меллером.
Классификация по генотипу:
1) Генные (точковые) мутации (Замена азотистого основания, Перекрестные замены, Вставка нуклеотида)
2) Хромосомные(оберации) мутации (дефишез, дереции, дубликация, Инверсия, транслакация, образование изохромосом)
3) Геномные мутации – связанные с изменением хромосом.
60. Репарирующие (исправляющие) системы клеток.
Репарация — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических агентов. Осуществляется специальными ферментными системами клетки. Ряд наследственных болезней связан с нарушениями систем репарации.
Возможна только в интерфазе, при репликазе (синтезе) ДНК. Восстанавливает не все повреждения, только те связанные с ультрофиолетом.
Репорации:
1) Тимидиновые тимберы- Световая или фотореактивация – происходит на свету, под влиянием специального фермента – фотореактиватора. Под влиянием ферментов связи тимедина разрушается, и восстанавливаются нормальные водородные связи.
2) Темновая репорация – вырезают фрагмент с тиминами, а потом он заново застраивается по принципу комплементарности .
3) Постреплекационный и рекомбинационный – происходит во время репликации ДНК, образуется новая цепочка ДНК, а 2 цепочка на разрушенном участке, потом по принципу комплементарности достраивается пустой фрагмент и появляется новая не поврежденная молекула ДНК.
61. Закон гомологических рядов в наследственной изменчивости Н.И. Вавилова.
1920- сформирован закон в Саратове.
Близкие виды и роды характеризуют сходные рядами в наследственной изменчивости с такой правильностью, что зная ряд форм в приделах 1-ого вида можно предвидеть нахождения параллельных форм у других видов и родов, чем ближе генетические виды и роды, тем полнее сходства ряда его изменчивости.
62. Множественный аллелизм.
Множественный аллелизм — это существование в популяции более двух аллелей данного гена. В популяции оказываются не два аллельных гена, а несколько. Множественный аллелизм для генов, контролирующих системы несовместимости, выступает как фактор отбора, препятствующий образованию зигот и организмов определенных зигот.
Примером множественного аллелизма является серия множественных аллелей s1, s2, s3, обеспечивающих самостерильность многих растений. Двенадцать различных состояний одного локуса у дрозофилы, обусловливающих разнообразие окраски глаз (w — белые, we — эозиновые, wa — абрикосовые, wch — вишневые, wm — пятнистые и т. д.); серия множественных аллелей окраски шерсти у кроликов («сплошная», гималайская, альбинос и т. д.); аллели IA, Iв, I°, определяющие группы крови у человека, и т. д.
Серия множественных аллелей — результат мутирования одного гена.
Обусловленность признака серий множественных аллелей не меняет соотношения фенотипов в гибридном потомстве. Во всех случаях в генотипе присутствует только одна пара аллелей, их взаимодействие и определяет развитие признака.
В результате мутирования возникает не два, а много аллелей одного гена.
Пример: Серия аллелей определяет окраску меха у кролика.
С(серая)˃  ˃
˃ 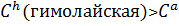 (альбинос)
(альбинос)
63. Геномные мутации, их особенности.
Геном – основной и гаплоидный набор хромосом. Геномные мутации связанны с изменением числа хромосом. Геномные мутации:
1. Автополиплоиды – это организмы, у которых хромосомный набор увеличен кратно гаплоидному набору.
п 2п 3п 4п ... 10 п ( у растений)
2. Гаплойды – одинарные набор хромосом. У трутней получают из не оплодотворенных яиц, у растений гаплоиды возможны, но мало жизнеспособны.
3. Анеуплоидия – в кариотипе на 1 или 2 хромосомы больше или меньше. Если меньше – аутосомы не жизнеспособны у животных, но у других жизнеспособны.
Если 2п -1 – моносомики(на 1 хромосому меньше)
У человека синдром Шерешевского в кариотипе 46 хромосом.
Если 2п-2 – нуллисомики – не жизнеспособные.
Если 2п + 1 – трисомики. Синдром Дауна, возможна лишняя у женщин Х-хромосома, у мужчин XY синдром Клайнфельтера.
Существует трисомия по 13 и 18 хромосом, но гибнут в эмбриогенезе.
Существует синдром Вай-вай XYY
64. Понятие о близкородственном и неродственном скрещивании.
Чистопородный метод разведения позволяет закрепить отличительные черты породы и быстро добиться однородности потомства. Этим методом добиваются постепен<
Последнее изменение этой страницы: 2016-08-11
lectmania.ru. Все права принадлежат авторам данных материалов. В случае нарушения авторского права напишите нам сюда...