Категории:
ДомЗдоровьеЗоологияИнформатикаИскусствоИскусствоКомпьютерыКулинарияМаркетингМатематикаМедицинаМенеджментОбразованиеПедагогикаПитомцыПрограммированиеПроизводствоПромышленностьПсихологияРазноеРелигияСоциологияСпортСтатистикаТранспортФизикаФилософияФинансыХимияХоббиЭкологияЭкономикаЭлектроника
РОЗДІЛ 2 МЕХАНІЗМ ТРАНСПОРТУ ГАЗІВ КРОВ’Ю ТА ЇХ ОБМІН У ТКАНИНАХ
Транспорт газів кров'ю
Кисень, що надходить у кров, спочатку розчиняється у плазмі крові. При РАО2 100 мм рт. ст. у 100 мл плазми розчиняється 0,3 мл О2.
Кисень, що розчинився у плазмі крові, за градієнтом концентрації проходить через мембрану еритроцита і утворює оксигемоглобін (НbО2). При цьому валентність заліза не змінюється. Оксигемоглобін — нестійка сполука і легко розкладається. Пряма реакція називається оксигенацією, а зворотний процес — дезоксигенацією гемоглобіну. При сполученні О2 із гемоглобіном Fe2+ залишається двовалентним [8].
Кожна молекула Hb може приєднати 4 молекули О2, що у перерахунку на 1 г Hb означає 1,34 мл О2. Знаючи кількість гемоглобіну в крові, можна визначити кисневу ємкість крові.
Враховуючи те, що 100 мл крові містять лише 0,3 мл розчиненого О2, можна уявити, що основний об'єм кисню транспортується у стані хімічного зв'язку з гемоглобіном. Але, незважаючи на відносно низьку розчинність, кількість розчиненого в крові О2можна збільшити штучно. Розчинність газу в рідині залежить від температури, складу рідини, тиску газу і його природи. Поза як склад крові, її температура у організмі майже завжди постійні, кількість розчиненого газу можна вирахувати за формулою. [11].
Коли збільшується тиск газу над рідиною, кількість розчиненого газу збільшується. Так, при диханні чистим О2, коли його парціальний тиск у альвеолах може перевищувати 600 мм рт. ст., у 100 мл крові розчиняється вже близько 2 мл кисню. Але якщо людина перебуває в умовах з підвищеним тиском кисню (у барокамері), то кількість розчиненого у крові кисню буде зростати пропорційно до тиску (гіпербарична оксигенація). Наприклад, при парціальному тиску 3 атм, коли РАО2 збільшується до 2280 мм рт. ст. (304 кПа), в 100 мл крові може розчинитись близько 5—6 мл О2. Цієї кількості кисню досить для того, щоб тканини не відчували кисневої недостатності навіть за відсутності зв'язаного з гемоглобіном О2. Вказаний ефект можна використовувати при наданні допомоги тим хворим, у яких гемоглобін не може транспортувати кисень. Наприклад, дихання чистим киснем рекомендована при отруєнні чадним газом, коли утворюється стійка сполука карбоксигемоглобін (дисоціює у 1000 разів повільніше, ніж оксигемоглобін).
Розчинність газів зменшується при підвищенні температури, але в умовах організму це великої ролі не відіграє. Про значення природи газу свідчить той факт, що розчинність кисню у 20—25 разів нижча, ніж вуглекислого газу [13].
Асоціація і дисоціація оксигемоглобіну
Між кількістю оксигемоглобіну в крові і напругою в ній кисню є залежність. Встановлено, що коли в крові немає кисню (Ро2 =0), то нема і оксигемоглобіну, він міститься у формі відновленого гемоглобіну. Якщо РО2 = 10 мм рт. ст., 5,5 % гемоглобіну перейде у форму НbО2, коли ж вміст Ро2 досягне 20 мм рт. ст., то оксигемоглобіну буде вже 25 %, при рівні 40 мм рт. ст. буде 74 %, а при рівні 100 мм рт. ст. у формі НbО2 буде близько 92 % О2. Таким чином, що більша напруга кисню в крові, то більше буде оксигемоглобіну.
Але ця залежність не лінійна, вона виражається S-подібною кривою. На рис. 2.1 показана залежність між РО2 крові і процентом НbО2 в ній.
Крива дисоціації оксигемоглобіну. Треба звернути увагу на дві частини цієї кривої. Верхня частина кривої, яка за формою наближається до горизонтальної, має значення для утворення оксигемоглобіну в капілярах легень. Сюда надходить венозна кров, у якій РО2 = 40 мм рт. ст. (в альвеолярному повітрі Ро2 =100 мм рт. ст.), тому кисень дуже швидко переходить у кров і рівень НbО2 зростає до 92 % [6].
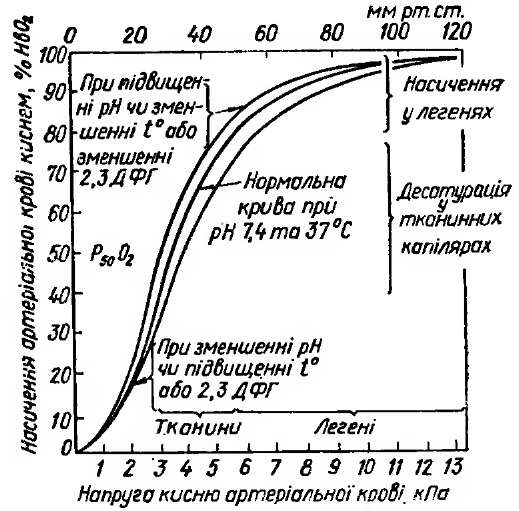
Рис. 2.1 - Криві дисоціації оксигемоглобіну
Але не завжди РО2 в альвеолярному повітрі буде становити 100 мм рт. ст. При підніманні на висоту, зниженні альвеолярної вентиляції він зменшиться. Проте, незважаючи на це, кількість гемоглобіну у формі НbО2 ще довго (до певної межі) буде досить високою і гемоглобін виконуватиме функцію транспорту кисню (цю частину кривої порівнюють із графічним зображенням процесу зарядження акумулятора).
Середня частина кривої, яка за формою наближається до вертикальної, має значення для процесів, що відбуваються в капілярах тканин. При РО2 40 мм рт. ст. (5,3 кПа) лише 72 % припадає на НbО2, але що інтенсивніше працюватиме орган, то менше буде в крові РО2 і більша буде дисоціація НbО2 на Hb і кисень. Отже, оксигенація клітин, що інтенсивно працюють, буде зростати. Цю частину кривої можна порівняти із графічним зображенням процесу розрядження акумулятора [9].
Показником, що характеризує інтенсивність використання 02 тканинами, є різниця кількості НbО2 в крові, яка притікає і відтікає,— артеріовенозна різниця (АВРО2).
Нахил кривої дисоціації оксигемоглобіну в крові людини не постійний і за деяких умов може змінюватися. Характер кривої дисоціації відображає показник, який має назву «напруга напівнасичення» (Р50). Р50 — така напруга О2, при котрій насичення гемоглобіну киснем складає 50 %. У нормі Р50 артеріальної крові становить близько 26 мм рт. ст. (3,5 кПа).
Конфігурація кривої дисоціації НbО2 обумовлена хімічною спорідненістю гемоглобіну до О2 й іншими зовнішніми факторами, що змінюють характер кривої. До таких факторів належать температура, pH, РСО2, концентрація в еритроциті 2,3-ДФГ [14].
Форма кривої дисоціації оксигемоглобіну значною мірою залежить від концентрації в крові Н+. При зниженні pH крива зміщується вправо, що свідчить про зменшення спорідненості Hb до О2. При підвищенні pH збільшується спорідненість Hb до О2 і крива зміщується вліво. Вплив pH на спорідненість Hb до О2 називається ефектом Бора. Ефект Бора відіграє певну роль у газотранспортній функції крові: утворення великої кількості СО2 в тканинах сприяє збільшенню віддачі кисню за рахунок зниження спорідненості Hb до нього. При виділенні СО2 у легенях зменшується pH крові і поліпшується оксигенація. СО2 також впливає на дисоціацію НbО2. Причому згаданий ефект обумовлений не тільки Н2СО3, але й прямим впливом рівня РСО2 на гемоглобін [6].
При зниженні температури віддача О2 оксигемоглобіном сповільнюється, а при її збільшенні прискорюється цей процес. Зміщенню кривої вправо сприяє також збільшення вмісту в еритроцитах 2,3-ДФГ. Вміст цієї речовини в еритроциті збільшується при анемії, що сприяє надходженню О2 до тканин і частково компенсує зниження рівня КЄК. Таким чином, відсутність у організмі запасів О2 компенсується за рахунок різкого збільшення використання його з крові, підвищення АВРО2. При інтенсивній роботі тканин, коли утворюється більше СО2, Н+ і підвищується температура, створюються умови для поліпшення доставки кисню клітинам.
У венозній крові міститься близько 580 мл/л СО2. У крові він міститься в трьох формах: зв'язаний у вигляді вугільної кислоти і її солей, зв'язаний з гемоглобіном і у розчиненому вигляді.
СО2 утворюється в тканинах при окислювальних процесах. У більшості тканин РСО2 складає 50—60 мм рт. ст. (6,7—8 кПа). У крові, що надходить у артеріальний кінець капілярів, РаСО2 становить близько 40 мм рт. ст. (5,3 кПа). Наявність градієнту змушує СО2 дифундувати із тканинної рідини до капілярів. Що активніше в тканинах здійснюються процеси окислення, то більше створюється СО2 і то більший Ртк.СО2. Інтенсивність окислення у різних тканинах різна. У венозній крові, що відтікає від тканини, PvСО2 наближається до 50 мм рт. ст. (6,7 кПа). А в крові, що відтікає від нирок, PvСО2 становить близько 43 мм рт. ст. Тому у змішаній венозній крові, що надходить до правого передсердя, у стані спокою PvСО2 дорівнює 46 мм рт. ст. (6,1 кПа) [4].
СО2 розчиняється в рідинах активніше, ніж О2. При PvСО2 який дорівнює 40 мм рт. ст. (5,3 кПа), у 100 мл крові розчинено 2,4— 2,5 мл СО2, що складає приблизно 5 % від загальної кількості газу, який транспортується кров'ю. Кров, що проходить через легені, віддає далеко не весь СО2. Велика частина його залишається в артеріальній крові, оскільки сполуки, які утворюються на основі СО2, беруть участь у підтриманні кислотно-основної рівноваги крові — одного із параметрів гомеостазу.
Хімічно зв'язаний СО2 перебуває в крові в одній із трьох форм: 1) вугільна кислота (Н2СО3): 2) бікарбонатний іон (НСО3); 3) карбогемоглобін (ННbСО2). У формі вугільної кислоти переноситься лише 7 % СО2, бікарбонатних іонів — 70 %, карбогемоглобіну — 23 %.
С02, який проникає у кров, спочатку піддається гідратації з утворенням вугільної кислоти: С02+Н2О  Н2СО3.
Н2СО3.
Ця реакція в плазмі крові відбувається повільно. В еритроциті, куди С02 проникає за градієнтом концентрації, завдяки спеціальному ферменту — карбоангідразі — цей процес прискорюється приблизно в 10 000 разів. Тому ця реакція відбувається в основному в еритроцитах (рис. 2.2). Утворювана тут вугільна кислота швидко дисоціює на Н+ і НСО3, чому сприяє постійне утворення вугільної кислоти: Н2С03  Н++НСО3.
Н++НСО3.

Рис. 2.2 - Транспорт СО2 кров'ю
У еритроциті при дисоціації вугільної кислоти Н+ утворює комплекс з відновленим гемоглобіном:
HbО2 О2 + Hb,
Hb + Н+ HHb [2].
При накопиченні НСО3 в еритроцитах створюється його градієнт з плазмою. Можливість виходу НСО3 у плазму визначається такими умовами: вихід НСО3 повинен супроводжуватися одночасним виходом катіону або ж надходженням іншого аніону. Мембрана еритроциту добре пропускає негативні, але погано — позитивні іони. Найчастіше утворення і вихід НСО3 із еритроцитів супроводжується надходженням у клітину Сl. Це переміщення називають хлоридним зрушенням.
У плазмі крові НСО3, взаємодіючи з катіонами, створює солі вугільної кислоти. У вигляді солей вугільної кислоти транспортується близько 510 мл/л СО2.
Крім того, СО2 може зв'язуватися з білками крові: частково — з білками плазми, але головним чином — з гемоглобіном еритроцитів. При цьому СО2 взаємодіє з білковою частиною гемоглобіну — глобіном. Гем же залишається вільним і зберігає здатність гемоглобіну перебувати одночасно у зв'язку як з СО2, так і з О2. Таким чином, одна молекула Hb може транспортувати обидва гази [5].
У крові альвеолярних капілярів усі процеси здійснюються в протилежному напрямі. Головна із хімічних реакцій — дегідратація — відбувається в еритроцитах за участю тієї ж карбоангідрази:
Н++НСО3 Н2СО3 Н2О + СО2.
Спрямування реакції визначається безперервним виходом СО2 з еритроциту у плазму, а із плазми в альвеоли. У легенях у зв'язку з постійним його виділенням відбувається реакція дисоціації карбогемоглобіну:
ННbСО2+О2 → ННbО2 + СО2 → НbО2 +Н++СО2.
Взаємозв'язок транспорту кисню та двооксиду вуглецю. Вище вказувалося, що форма кривої дисоціації оксигемоглобіну впливає на вміст СО2 у крові. Ця залежність пов'язана з тим, що дезоксигемоглобін є слабшою кислотою, ніж оксигемоглобін, і може приєднувати більше Н+. Унаслідок цього при зменшенні вмісту оксигемоглобіну підвищується ступінь дисоціації Н2СО3, а отже, збільшується транспорт СО2 кров'ю. Ця залежність називається ефектом Холдейна (рис. 2.3).
Взаємозв'язок обмінів двооксиду вуглецю і кисню яскраво виявляється в тканинах і легенях. До тканин надходить оксигенована кров. Тут під впливом СО2 посилюється дисоціація гемоглобіну. Тому надходження кисню в тканини сприяє прискоренню поглинання СО2 кров'ю [7].

Рис. 2.3 - Криві вмісту CO2 в оксигенованій (1) та дезоксигенованій (2) крові (за даними Р. Шмідта та Г. Тевса, 1986). Лінія «а — в» характеризує фактичний газообмін
У легенях відбуваються зворотні процеси. Надходження О2 знижує спорідненість крові до СО2 і полегшує дифузію СО2 в альвеоли. Це, в свою чергу, активізує асоціацію гемоглобіну з киснем [10].
Обмін газів у тканинах
На газообмін у тканинах впливають ті ж самі 5 факторів, що і на газообмін у легенях,— площа дифузії, градієнт напруги газів у крові і клітинах; відстань, яку проходить газ; коефіцієнт дифузії; стан мембран, через які проходять тази. Кількість О2, що надійшла до органа, може бути визначена на підставі об'єму кровообігу і вмісту О2 в приносній артерії і вені. АВРО2 і кровотік залежать від рівня метаболізму органа: що інтенсивніший обмін речовин, то більше використовується О2.
О2 використовується в мітохондріях клітин для реалізації процесу окислення. Тому найменше вільного О2 концентрується поряд з мітохондріями.
У нормі при найвищій активності окислювального процесу мінімальний Ро2 біля мітохондрій не повинен бути меншим від 1 мм рт. ст. Це критична напруга Ог в мітохондріях. При нижчому її показнику процес окислення стає неможливим. Р° біля мітохондрії становить 5—10 мм рт. ст. (0,7—1,3 кПа). У тканинній рідині біля капіляра Ро2 досягає 30 — 40 мм рт. ст. (4— 5,3 кПа), а у крові, яка надходить,— 100 мм рт. ст. Перепад Ро2 від крові капіляра до мітохондрій забезпечує процес дифузії О2 за градієнтом концентрації. Природно, що клітини, які прилягають до капіляра, забезпечуються киснем краще. Тому в клітині мітохондрії, які розташовані ближче до мембрани, також отримують більше кисню. Весь шлях, що проходить кисень від атмосферного повітря до мітохондрій клітин, нагадує ряд каскадів (рис. 2.4) [3].

Рис. 2.4 - Зміни РО2 на етапах дихання
При інтенсифікації споживання О2 рівень РО2 біля мітохондрій стає набагато нижчим від 10 мм рт. ст., що й сприяє прискоренню дифузії. У свою чергу зниження РО2 біля капіляра до рівня, що нижчий від 40 мм рт. ст. (5,3 кПа), різко стимулює дисоціацію оксигемоглобіну. Та частина кисню артеріальної крові, що застосовується тканинами, називається коефіцієнтом утилізації кисню. У стані спокою він дорівнює близько 30%, а при інтенсивній праці підвищується до 60—70 % (рис. 2.5) [1].

Рис. 2.5 - Дифузія О2 в тканинах
У протилежному напрямі від мітохондрій надходить утворений при окисленні СО2. Звичайно РСО2 у тканинах перебуває на рівні близько 60 мм рт. ст. (8,0 кПа), а біля капіляра — близько 50 мм рт. ст. (6,7 кПа). Тому СО2 надходить від мітохондрій до міжклітинної рідини, а звідти — до кровоносних капілярів і у кров.
ВИСНОВКИ
Таким чином, газообмін в легенях здійснюється по принципу дифузії. Кров у легенях постачається по судинах малого кола кровообігу. По верхній і нижній порожнистих венах венозна кров впадає у праве передсердя, а потім у правий шлуночок. Після чого кров з шлуночка виштовхується в легеневу артерію, що розгалужується до дрібних альвеолярних капілярів, де і проходить газообмін. Концентрація вуглекислого газу в крові більша, ніж в повітрі альвеол, а кисню концентрація в повітрі більша, ніж в крові, тому кисень переходить в кров, а вуглекислий газ – в повітря, яке під час вдиху виводиться в зовнішнє середовище.
Отже, в легенях кров збагачується киснем і звільняється від вуглекислого газу, тобто перетворюється із венозної у артеріальну. По легеневих венах уже артеріальна кров впадає в ліве передсердя, потім в лівий шлуночок, звідси – у велике коло кровообігу.
Із капілярів великого кола кровообігу кисень потрапляє в тканини. В артеріальній крові концентрація кисню більша, ніж в клітинах тканин, а вуглекислого газу в клітинах концентрація більша, ніж в крові. Тому кисень легко проникає в клітини тканин і використовується в окислювальних процесах. А вуглекислий газ із клітин переходить у кров. Так у тканинах органів артеріальна кров перетворюється на венозну.
Венозна кров по венах великого кола кровообігу надходить у праве передсердя, потім – у правий шлуночок серця, а звідти – до легень.
Кров, проходячи по легеневих капілярах, обмінюється газами з альвеомерним повітрям. У венозній крові, що надходить до легенів, високий тиск вуглекислого газу і відносно низький тиск кисню. У тканинах тиск вуглекислого газу найбільший, у венозній крові, яка надходить до альвеол, тиск дорівнює 46 мм, а в альвеолярному повітрі – 40 мм рт. ст. Завдяки такій різниці тиску вуглекислий газ дифундує з тканин у кров капілярів, а з венозної крові – в альвеолярне повітря.
Споживання тканинами кисню залежить від інтенсивності обміну і від функціонального стану тканин. На відміну від гідролітичних процесів, реакції окислення дають більший тепловий ефект і є для організму основним джерелом енергії.
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
1. Анатомия и физиология человека: Учебное пособие. Изд. 2-е. — Ростов н/Д: изд-во: «Феникс», 2003. - 416 с.
2. Біологія: Навч. посіб. / А. О. Слюсарєв, О. В. Самсонов, В.М.Мухін та ін.; За ред. та пер. з рос. В. О. Мотузного. — 3тє вид., випр. і допов. — К.: Вища шк., 2002. — 622 с.
3. Власова И.Т. Чеснокова С.Л. Регуляция функций организма: Физиологический справочник. - М, 1998. – 314 с.
4. Коробков А.В., Чеснокова СЛ. Атлас по нормальной физиологии. — М.: Высшая школа, 1986. – 428 с.
5. Максимов В.И. Основы анатомии и физиологии человека. М.: КолосС, 2004 – 167 с.
6. Основы физиологии. В 3-х тт. / Под ред. Б.И.Ткаченко. — СПб.: Международный фонд истории науки, 1994. – 411 с.
7. Основы физиологии человека: Учебник дня студентов вузов, обучающихся по медицинским и биологическим специальностям / Торшин, Владимир Иванонич, Власова Виктория Михайловна; Апджшо, Николай Александрович, 2-е издание, исправленное.- М.: РУДН, 2001. - 408 с.
8. Нормальна фізіологія / Кол. авторів; За ред. В. І. Філімонова.— К.: Здоров'я, 1994.— 608 с.
9. Нормальная физиология: учебник / Р. С. Орлов, А. Д. Ноздрачев. – М.: Гэотар-Медиа, 2005. – 696 с.
10. Нормальная физиология человека: учебник для высших учебных заведений / под ред. Б. И. Ткаченко. – М.: Медицина, 2005. – 928 с.
11. Физиология. Основы и функциональные системы: Курс лекций/Под ред. К.В. Судакова.— М.: Медицина, 2000.- 784 с.
12. Физиология человека. В 2-х тт. / Под ред. В.М.Покровского, Г Ф.Коротько. - М.: Медицина, 1997. – 539 с.
13. Физиология человека: В 3-х томах. Т. 2. Пер. с англ./Под ред. Р. Шмидта и Г. Тевса. — 3-е изд. — М.: Мир, 2005. — 314 с.
14. Физиология человека / Под ред. Н.А.Агаджаняна и В.И.Циркина. - Спб., 1998. – 290 с.
15. Физиология человека. Compendium: учебник для студентов высших учебных заведений / под ред. Б. И. Ткаченко и В. Ф. Пятина. – Самара: Самарский дом печати, 2002. – 416 с.
Последнее изменение этой страницы: 2016-07-22
lectmania.ru. Все права принадлежат авторам данных материалов. В случае нарушения авторского права напишите нам сюда...