Категории:
ДомЗдоровьеЗоологияИнформатикаИскусствоИскусствоКомпьютерыКулинарияМаркетингМатематикаМедицинаМенеджментОбразованиеПедагогикаПитомцыПрограммированиеПроизводствоПромышленностьПсихологияРазноеРелигияСоциологияСпортСтатистикаТранспортФизикаФилософияФинансыХимияХоббиЭкологияЭкономикаЭлектроника
РАЗДЕЛ II. ОБМЕН ВЕЩЕСТВ В ЖИВОМ ОРГАНИЗМЕ
ГЛАВА 3. БИОСИНТЕЗ НУКЛЕИНОВЫХ
КИСЛОТ И БЕЛКОВ
БИОСИНТЕЗ ДНК (РЕПЛИКАЦИЯ)
Процесс биосинтеза дочерней ДНК на ее матрице, приводяций к ее удвоению носит название репликации. Для реализации этого процесса необходима матрица — расплетенная цепь ДНК, субстраты, участвующие в полимеризации ДНК, ферменты, катализирующие этот процесс, ионы Mg2+, а также ряд белковых факторов, обеспечивающих деспирализацию двухнитевой ДНК.
Как и любой процесс, репликация протекает в несколько этапов: инициация, элонгация и терминация репликации.
Инициация репликации ДНК.
Репликация всегда предшествует делению клетки и начинается с расплетения двойной спирали ДНК. Это осуществляется при помощи ферментов хеликаз, которые перемещаются вдоль цепей ДНК и раскручивают их. Процесс расплетения спиралей ДНК является энергозависимым и требует затраты АТФ. Интенсивное раскручивание ДНК может привести к образованию дополнительных витков (сверхспиралей), которые легко предотвращаются ферментами топоизомеразами. Синтез новой цепи ДНК осуществляется при помощи ДНК-полимераз.
Элонгация репликации ДНК.
На следующем этапе репликации от 3'-конца цепи ДНК начинается синтез новой цепи нуклеиновой кислоты при помощи ДНК-полимеразы III. Синтез идет в направлении 5'®3' одновременно на обеих цепях матрицы. Поскольку цепи молекулы ДНК антипараллельны, только одна из новых цепей синтезируется непрерывно - эта цепь называется лидирующей. Цепь, направление синтеза которой противоположно движению репликативной вилки, называют отстающей, и синтез этой цепи имеет прерывистый характер (рисунок 3.1).
В процессе репликации на отстающей цепи образуется множество короткоцепочечных фрагментов, которые носят название фрагментов Оказаки. Их величина у прокариот составляет около 1000 нуклеотидов, у эукариот — в три раза меньше.
Терминация репликации.
Заключительной фазой процесса репликации является стадия терминации, или окончания синтеза. У эукарит, как и у прокариот, имеются специальные терминаторы, прекращающие синтез цепи ДНК. Этими терминаторами являются определенные последовательности нуклеотидов, при достижении которых ДНК-полимераза прекращается синтез новой цепи ДНК.
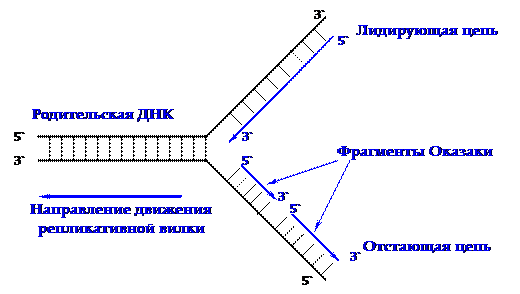 |
Рисунок 3.1 - Схематическое изображение репликативной вилки
Механизмы действия ДНК-полимераз эукариот и прокариот схожи между собой. Отличия в процессе репликации заключаются в следующем: хромосома эукариот имеет линейную структуру, на обеих цепях расположено множество репликонов и соответствующее количество терминаторов. Линейность ДНК эукариот является причиной проблем, которых не существует у прокариот, имеющих кольцевую ДНК. В отличие от лидирующей цепи, которая реплицируется полностью, праймер, находящийся у 3'-конца отстающей цепи, разрушается и не реплицируется при помощи ДНК-полимераз. Для предотвращения укорачивания цепи на концах хромосомы находятся теломеры — участки нереплицируемой ДНК. На этом участке ДНК может синтезироваться праймер, и полнота репликации сохранится. Теломера состоит из большого числа повторов, например у человека: ТТАГГГ.
БИОСИНТЕЗ РНК (ТРАНСКРИПЦИЯ)
Процесс биосинтеза РНК па матрице ДНК получил название транскрипции. Субстратами и источниками энергии для синтеза всех видов РНК являются рибонуклеозидтрифосфаты: АТФ, ГТФ, ЦТФ, УТФ. Катализируют синтез РНК ферменты РНК-полимеразы. В ядрах эукариотов обнаружены три специализированные РНК-полимеразы: РНК-полимераза I, синтезирующая пре-рРНК; РНК-полимераза II, ответственная за синтез пре-мРНК; РНК-полимераза III, синтезирующая пре-тРНК.
В процессе транскрипции, как и в репликации, различают три стадии: инициацию, элонгацию и терминацию. Кроме того, сюда можно отнести постранскрипционные изменения РНК
Инициация транскрипции РНК.
Инициация транскрипции является наиважнейшим фактором, определяющим начало синтеза РНК, его скорость и регуляцию.
Присоединение РНК-полимеразы к промотору определяет рамку считывания информации с матрицы ДНК. Активация промотора происходит с помощью белкового фактора (ТАТА-фактора), который облегчает взаимодействие промотора с РНК-полимеразой. Присоединение РНК-полимеразы к промотору увеличивает сродство фермента к факторам инициации (А, В), которые инициируют раскручивание примерно одного витка двойной спирали ДНК.
Далее к РНК полимеразе присоединяется кофермент, и образуется закрытый транскрипционный комплекс. В результате раскручивания цепей ДНК разрываются водородные связи между парами нуклеотидов, и образуется открытый транскрипционный комплекс.
Сразу после начала транскрипции первый нуклеотид с 5'-конца подвергается модификации (метилированию гуанина), которая называется кэпированием (от англ. «сар» - кепка). Поэтому первый нуклеотид, стартовая точка гена, называется кэп-сайт. Сигнал ААТААА (последняя последовательность, кодируемая РНК-полимеразой) способствует блокированию 3'-конца мРНК.
Элонгация транскрипции РНК.
Факторы элонгации (Е, Н, F) повышают активность РНК-полимеразы и облегчают локальное расхождение нуклеотидных цепей. Синтез молекулы РНК идет по направлению от 5`-конца к 3`-концу комплементарной матричной цепи ДНК.
После образования нескольких пар оснований происходит отделение РНК-полимеразы от транскрипционного комплекса, а водородные связи между нуклеотидами матрицы (цепь ДНК) вновь восстанавливаются. У прокариот частично синтезированная мРНК уже взаимодействует с рибосомами и вовлекается в процесс синтеза белка. В клетках эукариот синтез РНК и белка разобщен, кроме того, новосинтезированные транскрипты подвергаются посттранскрипционным модификациям.
Терминация транскрипции РНК.
Терминация синтеза РНК у прокариот обусловлена наличием на матрице таких последовательностей, которые допускают возможность образования шпильки на транскрипте РНК. При этом связь транскрипта с матрицей значительно ослабляется, что в конечном счете приводит к отделению РНК.
Механизмы терминации транскрипции у эукариот до конца не изучены. Полагают, что с РНК-полимеразой взаимодействует белковый стоп-сигнал, который замедляет (но не прекращает) транскрипцию. Далее фермент катализирует синтез последовательности ААУААА и следующие за ней 15 нуклеотидов, после чего завершает свою работу. В процессе отделения транскрипта от матрицы экзонуклеаза отщепляет терминальные 15 нуклеотидов, а фермент полиА-полимераза достраивает к последовательности ААУААА порядка 150—200 полиадениловых нуклеотидов (полиА).
Последнее изменение этой страницы: 2017-07-16
lectmania.ru. Все права принадлежат авторам данных материалов. В случае нарушения авторского права напишите нам сюда...