Категории:
ДомЗдоровьеЗоологияИнформатикаИскусствоИскусствоКомпьютерыКулинарияМаркетингМатематикаМедицинаМенеджментОбразованиеПедагогикаПитомцыПрограммированиеПроизводствоПромышленностьПсихологияРазноеРелигияСоциологияСпортСтатистикаТранспортФизикаФилософияФинансыХимияХоббиЭкологияЭкономикаЭлектроника
Эволюционное моделирование. Проблемы формы и содержания
Эволюционное моделирование. Проблемы формы и содержания
Новости искусственного интеллекта №5, 2003
Введение
Сложно сказать точно, когда впервые возникла простая идея, заключающаяся в том, что вместо моделирования сложного объекта в уже готовом виде следует заниматься моделированием эволюции некоторого простого объекта. Действительно, запустить эволюционный процесс и ждать, когда с течением времени появится требуемый ответ – это достаточно естественный и привлекательный путь решения сложно формализуемых задач.
История развития этого направления, называемого эволюционным моделированием, весьма характерна и показательна. Не успев оформиться в некоторую строгую и формальную теорию, эволюционное моделирование (ЭМ) на некоторое время уходит с арены активно разрабатываемых методов искусственного интеллекта. Затем, то ли по причине кризиса идей в ИИ, то ли по непреложному закону переоткрытия уже известного, но термин ЭМ спустя много лет возникает вновь. Правда, уже как новое направление ИИ, объединяющее в себе множество методов и моделей со своей классификацией и всеми вытекающими из узкой специализации последствиями.
Да и само понимание методологии ЭМ претерпело такие метаморфозы, что была утеряна изначальная цель этого направления. Дело не в формальных определениях и терминологии. Дело в том, что сложившееся понимание ЭМ перестало быть конструктивным.
В этой статье мы попытаемся обосновать необходимость возврата к пониманию сути ЭМ в изначальном его смысле, что должно привести к появлению предпосылок для создания целостной, единой методологии и принципов построения эволюционных интеллектуальных систем.
История вопроса
Впервые, пожалуй, идея ЭМ была явно сформулирована еще в 60-х гг. Л.Фогелем в работе «Искусственный интеллект и эволюционное моделирование» ([Fogel, 1966] или в русском переводе [Фогель Л. и др., 1969]), которую можно было бы назвать классической, если б она не была, фактически, предана незаслуженному забвению. Причин тому можно выделить несколько.
Начнем с того, что работы Фогеля появились в крайне неудачное для них время. То было время бурного расцвета ИИ, когда одна за другой появлялись новые концепции создания интеллектуальных систем, когда казалось, что вот-вот появится некая панацея, и искусственный интеллект, как синоним искусственного разума, будет создан. Поэтому идея эволюционного моделирования была лишь одной в ряду многих.
Во-вторых, сама работа во многих отношениях не выдерживала критики. К сожалению, авторы лишь декларировали основной эволюционный принцип создания искусственного интеллекта. С содержательной же точки зрения была выбрана далеко не лучшая структура эволюционирующего объекта (в виде детерминированного конечного автомата). А главное, воплощение основного механизма – процесса эволюции – было абсолютно неудовлетворительным. Можно сказать, что процесс эволюции как таковой полностью выпал из рассмотрения.
Самое интересное, что параллельно, практически одновременно с работами Фогеля создавались разнообразные модели, которые могли бы стать основой для эволюции. Например, исследования М.Л.Цетлина по теории автоматов ([Цетлин, 1969]) должны бы дать эволюционному моделированию замечательную модель объекта эволюции. С другой стороны, интенсивно развивались и формализовывались модели эволюции как таковой ([Шмальгаузен, 1968]). Однако пересечения этих трех составляющих ЭМ – самой концепции создания сложных систем на основе эволюции простейших, модели эволюционирующей особи и формальной эволюционной теории – в тот момент, к сожалению, не произошло.
Когда стало ясно, что создание искусственного разума откладывается на неопределенный срок, когда говорить о том, что ИИ и искусственный разум являются синонимами, стало неприличным проявлением дилетантизма (вопросы о том, нужно ли вообще воплощать все проявления человеческого разума, чем отличается интеллект от разума и т.п. мы рассматривать не будем), когда оформился широкий ассортимент узких специализаций направлений в области ИИ, тогда началась постепенная реинкарнация основного тезиса ЭМ о необходимости моделирования процесса эволюции.
Ознаменовалось это прежде всего появлением такого явления, как генетические алгоритмы. Предложенные Холландом [Holland, 1975] генетические алгоритмы (ГА), как водится, изначально претендовали на революционный метод решения интеллектуальных поисковых задач. Приверженцы этого направления утверждают, что ГА основываются на теории эволюции Дарвина (а именно, на идее естественного отбора). О проблемах ГА мы поговорим позже. Пока же достаточно сказать, что сейчас ГА рассматриваются значительно более скромно – лишь как некий стохастический оптимизационный метод, иногда дающий неплохие результаты. Действительно, находить экстремумы с помощью ГА можно. Но не более.
В то же время предпринимались попытки возобновления исследований в области ЭМ. Речь идет прежде всего о работах И.Л.Букатовой ([Букатова, 1975]). Однако даже в одной из наиболее значительных работ ([Букатова и др., 1991]) факторы эволюции лишь декларируются. Фактически речь идет о повторении исследований Фогеля на несколько более высоком качественном уровне с улучшенной методологической (декларативно) проработкой. Даже основной объект эволюции остался на том же уровне, что и у основателей ЭМ.
Заканчивая предысторию вопроса, можно сделать еще пару замечаний. Не исключено, что само мышление человека (по крайне мере та часть, которая относится к понятию инсайта – озарения) является эволюционным процессом. Речь идет о переборе на подсознательном уровне вариантов решения задачи [Тихомиров, 1966]. Раз перебор – значит, необходим механизм эффективного решения этой комбинаторной задачи. А это и есть предмет ЭМ. Кроме того, принятый в ЭМ подход достаточно хорошо сходится и с представлениями биологов относительно того, что отличает живые организмы высокого уровня развития от примитивных. Например, Р.Фокс в [Фокс, 1992] пишет: «биологическое преимущество развитой нервной системы состоит в способности к предсказанию нелинейных событий на основе быстрого моделирования». Тот же Фокс приводит примеры того, как осуществляется это быстрое прогнозирование в системах двигательного контроля, а также в зрительной коре хордовых.
Современность
Говоря о современном состоянии вопроса, главное – не потонуть в огромном количестве терминов, общим для которых является добавка «эволюционное»: эволюционная кибернетика, эволюционная биокибернетика, эволюционные модели и прочие эволюционные, желательно, многоагентные системы. К эволюционному моделированию относят и т.н. модели искусственной жизни (эволюционные, конечно), и молекулярно-генетические системы (вплоть до модели ДНК Уотсона и Крика), т.е. все, в чем так или иначе может фигурировать эволюция, биология и т.п.
Много говорят о генетических алгоритмах, о более экзотических моделях (например, о т.н. «сайзерах», которые, если отбросить биологическую терминологию, на поверку оказываются вариациями на тему фоннеймановских самовоспроизводящихся автоматов) и т.п., а также о моделях искусственной жизни – в лучшем случае – подобиях работ М.М.Бонгарда, изобилующими, в отличие от последних, массой спекулятивных рассуждений на околобиологические темы.
Весьма показательными в этом смысле являются работы Кауффмана, в которых исследуются аспекты «биологической эволюции» на основе т.н. NK-автоматов, состоящих из множества случайно связанных между собой логических элементов. Говорят, что NK-автоматы – это модель генетической регуляторной системы клеток.
Методология ЭМ
Оставаясь в рамках декларировавшихся изначально целей ЭМ, рассмотрим следующий вопрос. Возможен ли в ЭМ переход от задачи поиска экстремума в заданном пространстве задач (структур, состояний и т.п.) к задаче формирования качественно новых, сложных, выходящих за пределы начального пространства объектов. Далее будет дана попытка получения конструктивного ответа на этот вопрос.
Начнем с того, что эволюционное моделирование, как метод решения задач, можно представить в виде тройки
ЭМ = <М, О, З>
где М – модель эволюции, О – эволюционирующий объект, З – решаемая задача или критерии эволюции.
Далее мы рассмотрим все эти составляющие ЭМ, стараясь минимизировать конъюнктурные соображения. Вообще же одним из основополагающих тезисов эволюционного моделирования является естественность как реализации процесса эволюции и его механизмов, так и формы представления решения.
Модель эволюции
Положим в основу модели эволюции некоторую аксиоматику, задающую необходимые и достаточные условия, которые определяют ход эволюции: 1) наследственная изменчивость, т.е. мутирование, как предпосылка эволюции, ее материал; 2) борьба за существование как контролирующий и направляющий фактор; 3) естественный отбор как преобразующий фактор ([Шмальгаузен, 1968]).
Считается, что это – модель дарвиновской эволюции (хотя, справедливости ради, заметим, что ни в 1859 году в своем «Происхождении видов», ни в «Происхождении человека», опубликованном 12 годами спустя, Ч.Дарвин подобным образом эволюцию не определял).
Отметим еще раз, что автор ни коим образом не пытается дать свое толкование принципам эволюции. Более того, автор готов легко согласиться с любыми аргументами как в пользу теории дарвиновской эволюции, так и против нее. Тем более что, судя по всему, проблем в теории эволюции накопилось немало.
Приведем лишь некоторые «впечатляющие» тезисы противников теории естественного отбора.
· Геном человека лишь наполовину сложнее генома дождевого червя, а геном древесной лягушки в сотни раз больше генома «царя природы».
· Теория эволюции никак не объясняет миллионы лет застоя, чередующиеся с бурным образованием видов.
· С помощью экспериментов можно подтвердить лишь внутривидовые изменения, но нельзя объяснить происхождение новых видов. Да и между видами нет, якобы, переходных форм.
· Противоречит принципу естественного отбора тот факт, что некоторые структуры организмов не несут никакой физиологической нагрузки, а ранние стадии эволюции вообще никак не связаны с адаптивной функцией.
· Нет надежных теорий, объясняющих появление и наследование вариаций. Неясен механизм наследования жизненных изменений.
· Нет объяснений росту организма, развертыванию внутренне заложенного в нем плана, а принципом отбора никак не объяснить появление неадаптивных черт и вымирание.
· В рамках естественного отбора плохо объясняется связь поведения животных с генетической ассимиляций.
Более того, некоторыми исследователями считается, что биогеоценоз появился изначально как целостная, единая система, а на практике мы наблюдаем не происхождение новых видов, а вымирание целых популяций животного мира (об этом говорил еще Платон, рассуждая о человеке и животном мире).
И, наконец, вместо теории эволюции предлагают теорию периодических катастроф (исследования Шеклтона и Опдайка). Речь идет о периодическом - раз в сто тысяч лет – изменении температуры вод Мирового океана. При таком положении дел эволюция в столь короткие сроки невозможна.
А подытожить вышеизложенное можно универсальным патетическим заявлением: до сих пор наука не может достаточно внятно объяснить не только происхождение человека, но и хотя бы приблизительные сроки его появления на Земле. Каждое очередное палеонтологическое открытие становится сенсационным и «заставляет пересмотреть взгляды на эту проблему».
Все это приведено лишь для того, чтобы подчеркнуть понимание автором той ответственности, которую накладывает использование «эволюционных» терминов и механизмов в рассматриваемом сугубо формальном методе решения задач.
Итак, будем считать, что в основе ЭМ, как метода решения задач, лежат не некоторые конъюнктурные и «субъективно-интуитивные» соображения, а достаточно объективная, хорошо описывающая наблюдаемые явления жизни на Земле аксиоматика.
Модель особи
Говоря о важности выбора модели эволюционирующего объекта, имеется в виду не что иное, как извечный вопрос формы. С одной стороны, исходя из принципа универсальности и глобальности самого метода ЭМ, декларируется, что субъектом эволюции может быть что угодно. Однако здесь все не так просто. «Неудачная» модель может погубить все дело. Так произошло, например, с генетическими алгоритмами. Дело даже не в том, что эволюция не может происходить без учета фенотипической деятельности организма. Этот, возможно, не совсем очевидный факт будет нами обсуждаться позже. На поверхности лежит другое соображение. Интерес представляет поиск априори неизвестного алгоритма решения некоторой задачи. Следовательно, структура особи должна нести в себе как минимум описание этого искомого алгоритма (либо его фрагмента, если речь идет о получении решения в виде популяции). При этом желательно, чтобы форма этого представления была бы как можно более естественной. Если же мы будем говорить теперь именно об эволюции алгоритмов, то останется лишь выбрать, исходя из практических соображений, удобную и/или естественную форму представления.
Достаточно очевидной представляется следующая цепочка:
… ® Алгоритм ® Машина Тьюринга ® Автомат ® …
С практической точки зрения наиболее простой формой является конечный автомат (который можно рассматривать как вырожденный, упрощенный случай представления алгоритма). Тогда, разумеется, речь будет идти об эволюции в пространстве, скажем, распознавателей регулярных языков. Разумеется, с тем же успехом можно говорить и об эволюции различных грамматик, множеств предикатов и т.д.
Задачи или критерии эволюции
Под эволюцией в ЭМ будем понимать процесс образования особей, успешно решающих обусловленные средой задачи. На самом деле задача заключается в банальном выживании. То, насколько устойчива численность популяции, и определяет степень успеха эволюции. Постановка же задачи в ЭМ заключается прежде всего в формировании среды, к которой и должны приспосабливаться члены популяции. Естественно, что с практической точки зрения интерес представляют прежде всего популяции, состоящие из особей, решающих задачу приспособления/выживания «самостоятельно», вне их связи с другими членами популяции. Но, как будет видно в дальнейшем, иногда можно рассматривать эволюцию не особи, а популяции в целом (хотя, с точки зрения биологии, эволюция имеет интегральный, популяционный, видовой характер).
Итак, рассмотрим абсолютно абстрактную, не имеющую практически ничего общего с конъюнктурными (в хорошем смысле этого слова) представлениями об эволюции модель.
Реализация моделей
Сформулируем следующую задачу. Пусть дано множество примеров – конечных последовательностей пар символов некоторого алфавита. Необходимо построить модель, «объясняющую» некоторым образом эти примеры. Это – не что иное, как типичная задача индуктивного вывода. При этом пусть в качестве объясняющей модели выступает конечный автомат.
Решение будем искать следующим образом. Возьмем некоторое начальное множество автоматов произвольной структуры. Каждый из них независимо от остальных будет анализировать множество примеров. Если автомат распознает множество примеров без ошибок, то этот автомат и есть искомое решение. В случае ошибочного распознавания автомат наказывается, происходит накопление штрафных очков. По достижении некоторой предельно допустимой суммы штрафных очков автомат «умирает». При этом периодически автоматы размножаются простым делением. Полученная дочерняя особь является почти точной копией родителя. Отклонения в структуре отпрыска – это результат «малых» мутаций. Таким образом, можно считать, что решение поставленной задачи будет искаться в ходе эволюции некоторой популяции автоматов.
Опишем теперь этот процесс более формально. Начнем с описания простейшей модели популяции, в которой отсутствует непосредственное взаимодействие особей. Такая модель может быть названа «автоматным газом».
Строго говоря, автоматный газ представляет собой синхронно функционирующее в дискретные моменты времени множество конечных автоматов, генерирующих выходные сигналы в соответствии с некоторой стохастической матрицей. Классификация входного вектора осуществляется циклически. На каждом такте функционирования на вход автоматов подаются входные сигналы.
При этом вводится функция ошибки, устанавливающая соответствие между входной и выходной последовательностями - некий функционал вида
S(x1,x2,..,xn,y1,y2,...,yn),
где x1,x2,..,xn- входная последовательность, y1,y2,...,yn- выходная последовательность символов. Функция ошибки определяет величину штрафа, получаемого автоматом за неверный выходной сигнал (действие или предсказание). Когда величина накопленных особью штрафов превышает некоторое заданное значение, данная особь «умирает» (аналог естественного отбора). Автомат содержит стохастическую матрицу P, определяющую вероятности действий. Выходная реакция автомата yt определяется стохастическим вектором из P, соответствующим состоянию автомата и входному сигналу.

Особи периодически размножаются. Полученная в результате партеногенетического размножения особь наследует свойства (т.е. структуру) своего родителя. Однако при копировании структуры происходят локальные мутации, что обуславливает такой необходимый фактор эволюции, как наследственная изменчивость. Под локальными мутациями понимается множество элементарных структурных изменений: изменение количества состояний автомата, изменение связей между состояниями и т.п. В свою очередь, роль борьбы за существование играет конкуренция коэффициентов размножения автоматов в условиях ограниченного размера популяции.
В результате эволюции популяции происходит процесс формирования особей, которые с минимальными ошибками классифицируют входные последовательности. До сих пор считалось, что фактически мы имеем дело с нерегрессивным поиском в пространстве структур (см., например, [Букатова и др., 1991]). Однако, как будет показано ниже, эволюция – это поиск не только и не столько в пространстве структур (пространстве генотипов), но поиск в комбинированном пространстве – пространстве гено-фенотипов.
Рассмотрим теперь среду, в которую погружены эволюционирующие особи. Назовем некий набор элементарных признаков (ЭП) ji(Xi,Vi) составной средой C={j1,j2,..,jm}. ji- это двойка: вектор признаков Xi=(x1i,x2i,..,xni) и значение характеристической функции (номер класса, если речь идет о решении задачи классификации) Vi , Vi=1,..,n. Понятие составной среды C отличается от составной среды, введенной Цетлиным [Цетлин, 1969]. Различие это заключается прежде всего в детерминированном характере компонентов составной среды. Функционирование распознающей системы состоит в периодическом случайном выборе очередного элементарного признака ji из среды C и предъявления его множеству автоматов. Данный ЭП можно, в свою очередь, рассматривать как стационарную среду, в которую погружается автомат. Мы же, поскольку речь здесь идет о задаче индуктивного вывода, будем трактовать ЭП jiкак очередной пример. Индуктивный вывод будет делаться именно на основе множества примеров C. Итак, автомату A периодически предъявляются примеры среды C, правильность классификации которых служит мерой эффективности распознающего (классифицирующего) алгоритма (или автомата, что суть одно и то же). Будем говорить, что множество ЭП фиксированной и одинаковой размерности представляет собой составную среду (С-среду).
И, наконец, введем понятие t-момента времени - значения локального времени, под которым будем понимать порядковый номер текущего символа анализируемой среды. В отличие от абсолютного значения времени моделирования, t-момент времени характеризует именно текущий вектор признаков, т.е. текущую стационарную среду. Особенностью решающих автоматов является отсутствие выделенных конечных (терминальных) состояний. Любое состояние, вообще говоря, может считаться конечным и становится им по достижении конца входной последовательности. В силу отсутствия выделенных терминальных состояний, каждый переход генерирует выходной символ - значение решающей (или классифицирующей) функции в данный t-момент времени.
Создав таким образом популяцию автоматов и среду их обитания, можно запускать процесс эволюции. Визуально за ходом эволюции можно наблюдать по графику, по оси абсцисс которого откладывается время, а по оси ординат – значение показателей качества автоматов – отношение правильно предсказанных символов к общему количеству предсказаний.
 t
t
Рис.1. Фрагмент временной диаграммы процесса эволюции
Как видно из графика, в начале эволюции качество предсказаний далеко от оптимального. С течением времени в популяции появляются автоматы, более успешно решающие задачу - показатель качества достигает максимума. В дальнейшем эволюция может прекратиться. Это означает, что вымрут «неоптимальные» особи и останутся лишь те, кого не за что будет наказывать. В связи с этим более «устойчивым» показателем качества популяции является средний возраст особей. В «благополучной» популяции (как и в обществе) преобладают «долгожители». Впрочем, и об этом будет говориться ниже, отсутствие «естественной» смертности вовсе не является единственным вариантом процесса моделирования. Иногда можно и даже нужно иметь некий небольшой уровень смертности особей.
Отметим, что, строго говоря, видообразование в рассматриваемых моделях отсутствует. Формально определение вида, как основной единицы эволюционного процесса выглядит примерно так. Вид – это таксономическая систематическая единица, группа особей с общими морфофизиологическим, биохимическим и поведенческим признаками (морфолого-функциональный критерий), способная к взаимному скрещиванию (следовательно, имеющая сходный генетический аппарат).
С другой стороны, решение, получаемое в ходе моделирования эволюции, т.е. множество особей, решающих (по отдельности или всей совокупностью) поставленную задачу может рассматриваться как появление вида-индикатора, т.е. вида, служащего показателем особенностей среды какого-либо биоценоза или экосистемы.
Факторы эволюции
Приведенная выше простая схема уже дает богатый материал для рассуждений. Одно лишь варьирование параметров модели позволяет обнаруживать интереснейшие явления и биологические аналогии. Правда, существует опасность чрезмерного увлечения этими «биологизмами». Мы постараемся избегать неоправданного привлечения биологической терминологии и вообще «притягивания за уши» генетических механизмов к модели. Далее мы рассмотрим такие эволюционные факторы, как мутации, роль фенотипа и пресловутое наследование благоприобретенных признаков. Эти факторы являются весьма показательными для уяснения сути методологии ЭМ.
Мутации
В ЭМ под мутацией понимается случайное изменение структуры дочерней особи. Например, добавление или удаление состояния, перехода и т.п. структурные изменения, если эволюционирующей особью является автомат. При этом считается, что изменения эти должны быть «малы» - появляться не часто, не быть кратными (комплексными) и т.д. Дело в том, что интенсивные мутации приводят к тому, что система «идет в разнос» - поиск (блуждание) в пространстве состояний приобретает хаотический характер, проскакиваются близлежащие локальные экстремумы и процесс часто завершается полным вымиранием популяции.
С точки зрения популяционной генетики ([Фогель Ф., 1990]) различают следующие типы мутаций:
· геномные мутации (изменяется число хромосом);
· хромосомные мутации (изменение структуры хромосом); их можно обнаружить при микроскопическом исследовании;
· генные мутации – обнаруживаются только путем генетического анализа фенотипических изменений.
Тогда изменение количества состояний автомата можно считать геномными мутациями, а изменение связей – хромосомными или генными.
Если говорить о количественных параметрах, то при проведении экспериментов частоты мутаций (по крайней мере – соотношение их вероятностей) определялись заимствованными из биологии оценками ([Фогель Ф., 1990]):
геномные мутации: 5.67*10–4...2.7*10–5;
хромосомные мутации: 5*10–5...1.15*10–3.
Иными словами, частота хромосомных мутаций больше геномных примерно в 10 раз.
Кроме того, существуют и т.н. кодонные мутации, суть которых заключается в замещении данного основания другим определенным основанием, в результате которого одна аминокислота заменяется на другую. Они достаточно редки. Их частота оценивается в 2.5*10–9. С некоторой натяжкой можно считать, что аналогом кодонных мутаций могут быть изменения таких параметров, как период размножения, использование режима наследования признаков родительской особи и т.п.
О роли фенотипа
Менее всего автору хотелось бы услышать обвинения в некомпетентности за вторжение в такую сугубо «биологическую» область, как оценка роли фенотипа организма. Споры на тему его значимости, дебаты о промежуточных видах и т.п. пусть остаются уделом специалистов. Мы же рассмотрим этот вопрос исключительно с «технической» точки зрения.
Итак, пусть мы имеем процесс блуждания точки или множества точек в пространстве состояний. Задача этого блуждания заключается в нахождении некоторого экстремума (локального или глобального – не важно; хотя лучше, конечно, глобального). Блуждая, точки-особи делают пробные шаги – оставляют потомство, отстоящее, благодаря мутациям, на некоторое расстояние от родителя. Оценка качества особи, накопление штрафов и т.д. играют роль управляющего воздействия. Существует ли механизм, позволяющий, с одной стороны, избежать «застревания» точки в локальном экстремуме с одной стороны, и обеспечивающий достаточно «мелкий» шаг блуждания (чтобы не пропустить искомый глобальный экстремум) – с другой?
Иными словами, необходим «запас прочности», чтобы точка смогла бы доползти до соседнего экстремума. Состояние точки между экстремумами можно назвать промежуточным видом. Аналогия здесь абсолютно естественна. Вот лишь одна из классических иллюстраций – летающие рыбы. Сначала была просто рыба. С короткими плавниками. В конце концов появилась рыба летающая, с длинными плавниками. А где же промежуточная рыба, с плавниками, уже достаточно длинными, чтобы они мешали ей плавать, но еще короткими для того, чтобы летать? Очевидно, что такая «промежуточная» рыба нежизнеспособна, потомства оставить не может (не успеет, вымрет раньше) и так далее.
Та же картина наблюдается и тогда, когда мы рассматриваем эволюцию автоматов. Предположим, что имеется автомат со структурой A1. Требуемый автомат Ak может быть получен последовательным применением операторов мутаций M:
A1 ® A2 =M(A1) ® A3 =M(A2) … ® Ak =Mk(A1)
Если структуры автоматов A1 и Ak отвечают состояниям экстремумов, то «промежуточные виды» Ai могут иметь худшие показатели качества, т.е. быть нежизнеспособными.
Механизм, позволяющий особи с явно неоптимальным генотипом все-таки выжить и успеть оставить потомство, может быть основан на его фенотипической деятельности.
Суть в том, что конечное значение имеет не нахождение оптимальной точки в пространстве структур, а создание (или выведение) особи, обладающей оптимальным или субоптимальным поведением. В этом случае можно говорить о поиске в неком объединенном фено-генотипическом пространстве, в котором различающиеся структурно (генотипически) особи могут вести себя одинаково. Тогда «неудачный» генотип может некоторое время компенсироваться приемлемым фенотипическим поведением.
В этом случае мы действительно получаем наблюдаемое в биологии явление: изменения фенотипа, вызванные мутациями, образуют непрерывный ряд от вариантов с практически неизменными биологическими функциями к тем, которые проявляются только в неблагоприятных условиях [Фогель Ф., 1990].
Фенотипическое приспособление, основанное, скажем, на рассмотренных выше условно-рефлекторных принципах, вполне может обеспечить кратковременную жизнеспособность организма, несмотря на его «неудачную» наследственность.
Отказ от взаимно однозначного соответствия между поведением особи и ее генотипом является достаточно серьезным шагом. С одной стороны, мы получаем более гибкую схему, позволяющей одному и тому же генотипу по-разному проявлять себя в условиях изменяющейся среды. С другой – мы получаем тот самый «непрерывный ряд» вариантов и т.н. «малые», «локальные» мутации. Кроме того, все это хорошо согласуется с результатами из основной предметной области – биологии. Во-первых, существует такое явление, как фенокопия – воспроизведение генетически детерминированного признака под воздействием внешних факторов ([Фогель Ф., 1990], [Гольдшмидт, 1935]). Во-вторых, известно, что большинство мутаций безразлично для организма. Речь идет о пресловутой проблеме «молчащей» ДНК. На самом деле, общая интенсивность мутаций весьма высока. Если речь идет о человеке, то при размножении происходит в среднем 17.5 мутаций на 1 половую клетку и 35 мутаций на индивида (вероятность замещения одного основания/аминокислоты на другое). Введение фено-генотипического пространства и оценки особи именно с точки зрения ее поведения позволяет достаточно естественно интерпретировать эти вопросы. Действительно, результаты большинства мутаций «нивелируются» фенотипической составляющей. Кроме того, «молчащие» (в конкретных условиях среды) фрагменты ДНК могут отвечать за постепенное «депонирование», накапливание новых видовых признаков.
Эволюция вида и эволюция особи
Одним из параметров моделирования является период размножения особи – количество тактов, по истечении которого автомат осуществлял деление. Обычно этот параметр был фиксированным. Однако стоило включить его в список возможных мутаций (строго говоря, в биологии это называется использованием в качестве наследуемой единицы фазы клеточного деления), как проявился один достаточно интересный эффект.
Дело в том, что при этом в ряде экспериментов популяция автоматов начинала вырождаться в совокупность примитивных по своей структуре особей, весьма неэффективно осуществляющих предсказания. Однако в целом популяция вела себя достаточно устойчиво за счет возросшего коэффициента размножения. Этакая совокупность короткоживущих, но крайне интенсивно размножающихся «идиотов». Этот момент представляется, вообще говоря, достаточно важным для того, чтобы остановиться на нем более подробно. Во-первых, подобное решение (высокая скорость размножения при неоптимальной, крайне упрощенной структуре особи) - это тоже оптимальное приспособление. Амеба приспособлена не хуже человека. Развитие простейших – такой же биологический прогресс, но только с точки зрения процветания вида [Горбань, 1988]. Во-вторых, возможно, в этом усматривается некий общий эволюционный принцип: при использовании в качестве наследуемой и мутируемой единицы периода размножения популяция имеет тенденцию не к морфологическому, а к биологическому прогрессу. В биологии тому есть немало примеров.
Как бы то ни было, нас интересует, конечно, именно морфологический прогресс, связанный с совершенствованием организма.
В качестве иллюстрации на Рис.3 приведены фрагменты графиков обобщенных показателей эволюции. При этом параметр «период размножения» (фаза клеточного деления) является мутируемым. Обратите внимание на то, что усредненный показатель качества sQu популяции близок к 1, средний возраст sAge – мал, а средний период размножения sTp в конце концов так же установился на очень небольшом уровне. Как видно, вариабельность фазы клеточного деления приводит, во-первых, к уменьшению периода размножения и, во-вторых, к упрощению структуры особей, что влечет за собой ухудшение показателя качества отдельной особи, но не популяции в целом.
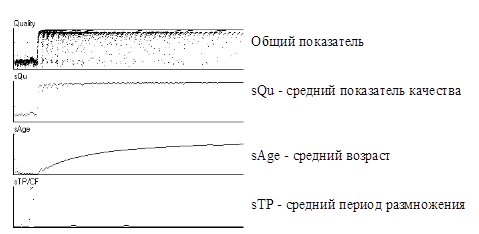
Рис.3. Эволюция вида
Синтезогенез
К.М. Завадским [Завадский, 1970] было введено понятие синтезогенеза - одного из путей эволюции, при котором происходит объединение простых организмов в более сложный. Примером синтезогенеза в природе являются переход от одноклеточных водорослей к многоклеточным, сообщество пчел в улье, муравьев в муравейнике и т.п. Однако это не просто скопление однородных подсистем. Совокупность элементов лишь тогда образует единую систему, когда эти элементы обладают потенциальным свойством образовывать статические или динамические структуры, необходимые для «выживания» элементов и всей их совокупности, т.е. обладают свойством устанавливать взаимодействие друг с другом для достижения локальных или глобальных целей.
Одним из примеров организации синтезогенетических систем является моделирование поведения автоматов на тороидальной поверхности [Варшавский, Поспелов, 1984]. По поверхности тора, разбитой на клетки, перемещаются автоматы с одним действием. При попадании в одну клетку нескольких особей происходит объединение их структур. Тем самым усложняется структура «организма» и, соответственно, изменяется манера его поведения. Следует отметить, что здесь мы имеем не реализацию мажоритарной логики напрямую - в составном объекте четко выражены составляющие его «одноклеточные». Поведение составного автомата строится на основе переключения между поведенческими актами образующих его элементарных автоматов.
С точки зрения эволюционного моделирования наиболее приемлемой формой синтезогенеза могла бы быть популяция из взаимодействующих особей, образующих сложные, возможно, иерархические структуры наподобие сообщества муравьев. Речь здесь идет об изучении моделей коллективного поведения и взаимодействия популяций однородных особей.
В основе теории коллективного поведения лежит гипотеза простоты, высказанная М.Л.Цетлиным [Цетлин, 1969]. Суть ее сводится к тому, что любое достаточно сложное поведение слагается из совокупности простых поведенческих актов. Их совместная реализация и простейшее взаимодействие приводят в результате к весьма сложным поведенческим процессам. Отсюда возникла идея о том, что совместное функционирование простых «маленьких зверушек» в сложной среде способно обеспечить устойчивое существование всего коллектива, который можно рассматривать как некий «сверхорганизм».
Выше неоднократно подчеркивалось, что особый интерес представляет морфологический прогресс, приводящий к созданию сложной структуры особи. Однако можно предположить, что функционально-логически между муравейником и, скажем, каким-нибудь «большим животным» различие не так уж велико. Здесь вновь возникает проблема формы-содержания, т.к. речь идет о логической модели с одной стороны и «физической», «пространственной» локализации частей организма – с другой. Совокупность отдельных «узкоспециализированных» особей эквивалентна единому сложному организму, состоящему из множества функциональных частей. Именно в этом смысле колония насекомых эквивалентна (функционально, с точки зрения умения решать задачи) единому, морфологически сложному организму.
Здесь мы подходим к самому проблемному моменту ЭМ – образованию сложных синтезогенетических структур. Схематично процесс функционирования, скажем, автоматного газа можно представить <
Последнее изменение этой страницы: 2016-07-22
lectmania.ru. Все права принадлежат авторам данных материалов. В случае нарушения авторского права напишите нам сюда...