Категории:
ДомЗдоровьеЗоологияИнформатикаИскусствоИскусствоКомпьютерыКулинарияМаркетингМатематикаМедицинаМенеджментОбразованиеПедагогикаПитомцыПрограммированиеПроизводствоПромышленностьПсихологияРазноеРелигияСоциологияСпортСтатистикаТранспортФизикаФилософияФинансыХимияХоббиЭкологияЭкономикаЭлектроника
Электрическая активность коры головного мозга у животных в состоянии голода и после приема пищи
Для того, чтобы характеризовать, каким образом возбуждения подкорковых отделов пищевого центра выходят на «кору мозга, в первой серии экспериментов нами изучался характер электрической активности коры мозга у животных в различные сроки голодания и после приема пищи.
Опыты-показали, что у подавляющего числа животных под уретановым наркозом после 1—2-суточного голодания в передних отделах коры головного мозга регистрировалась высокочастотная низкоамплитудная электрическая активность (амплитуда колебаний 6—10 мкв, частота 20—25 кол/сек), которая резко отличалась от высокоамллитудной (50—80 мкв), относительно медленной (6—8 кол/сек) активности теменно-затылочных областей.
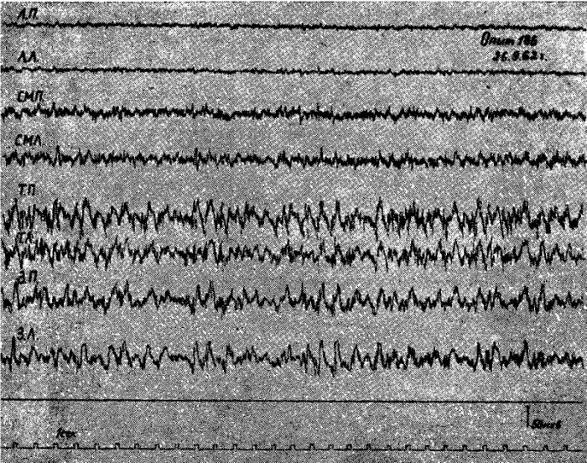
Рис. 1. Электрическая активность коры мозга кошки под уретановым наркозом после 2-суточного голодания.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сейсомоторная правая, CMЛ — сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек.
Граница распространения но жоре головного мозга высокочастотной активности у животных после 1—й-суточного голодания была определена в специальных опытах. Такая активность в наиболее отчетливой форме наблюдалась над отделами мозга, расположенными впереди венечной борозды. Однако высокочастотная активность регистрировалась и в каудальных частях передних отделов мозга, в которых по мере продвижения в направлении теменных отделов постепенно нарастала высокоамплитудная активность. Задней границей распространения зоны высокочастотной низкоамплитудной ЭЭГ активности у животных после кратковременного голодания была область сильвиевых борозд.
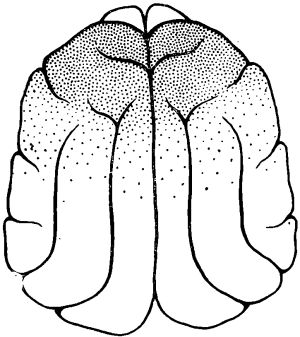
Рис. 2. Схема распространения под уретановым наркозом реакции, десинхронизации по коре мозга у кошки после 2-суточного голодания.
У накормленных перед опытом животных характер электрической активности коры мозга значительно отличался от активности голодных животных. У таких животных со всех отделах коры мозга регистрировалась медленная высокоамплитудная электрическая активность (амплитуда 50—80 мчсв, частота 3—5 кол/сек)
Все эти эксперименты указывали на то, что у животных в состоянии физиологического голода наблюдается ЭЭГ активация коры мозга. Эта активация под уретановым наркозом наиболее отчетливо проявляется в передних отделах коры мозга.
Наличие реакции десинхронизации корковой электрической активности передних отделов мозга у голодных животных указывало ка то, что эти отделы мозга находились у них в -состоянии возбуждения. При насыщении животного это возбуждение не наблюдалось. Естественно было предположить, что ЭЭГ активация передних отделов шоры мозга у голодных животных определялась «голодным» возбуждением пищевого центра.
Для выяснения того, в какой степени избирательная ЭЭГ активация передних отделов коры мозга у голодных животных была связана с деятельностью пищевого центра, мы провели специальные эксперименты.
Прежде всего мы исследовали, как скажется на ЭЭГ у голодных животных их искусственное насыщение.
Такое «насыщение» осуществлялось двояким путем: в одних случаях вводили в кровь голодного животного, находящегося под уретановым наркозом, раствор глюкозы или >кровь накормленного животного; в других опытах — животному под уретановым наркозом вводили жидкую пищу в ротовую полость, а затем в желудок.
При наличии у голодного животного реакции ЭЭГ активации только в передних отделах коры мозга и медленной высокоамллитудной активности в теменных и затылочных отделах, (введение в кровь глюкозы вызывало через 0,5—2 минуты значительное возрастание амплитуды и снижение частоты колебаний в передних отделах коры мозга. При этом иногда также увеличивалась амплитуда и снижалась частота колебаний в теменных и затылочных отделах коры мозга.

Рис. 3. Электрическая активность коры мозга кошки под уретановым наркозом после кормления.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сензомоторная правая СМЛ — сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек.

Pис. 4. Изменения электрической активности коры мозга у голодной кошки после введения внутривенно 40% — 2 мл глюкозы. Момент введения глюкозы обозначен стрелками. Обозначения: ЛП — лобная правая. ЛЛ — лобная левая. СМП — сензомоторная правая. СМЛ — сензомоторная левая. ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени I сек.
Следовательно, голодное состояние животного в какой-то степени влияло на деятельность и этих отделов коры.
Как правило, эффект введения глюкозы продолжался не более 15—20 минут, после чего электрическая активность коры мозга возвращалась к исходному активированному состоянию.
Аналогичные изменения ЭЭГ наблюдались при введении голодным животным крови накормленного животного.
С целью контроля в ряде опытов животным внутривенно вводили физиологический раствор. Однако введение физиологического раствора не изменяло заметно ЭЭГ голодного животного-
Произведенные опыты совершенно определенно указывали на значение гуморальных факторов для активации -коры мозга у голодных животных. Уже на основании этих опытов можно было считать, что активация передних отделов коры мозга у голодных животных определялась возбужденным «голодной» кровью пищевым центром.
Однако для более убедительного доказательства этого положения необходимо было исследовать, как изменится ЭЭГ картина голодного животного после натурального и естественного раздражения пищей.
Проведенные в этом направлении эксперименты доказали, что орошение рецепторов ротовой полости молоком, также как и введение молока в желудок, в ряде опытов приводило к временному устранению активации ЭЭГ у голодных животных.
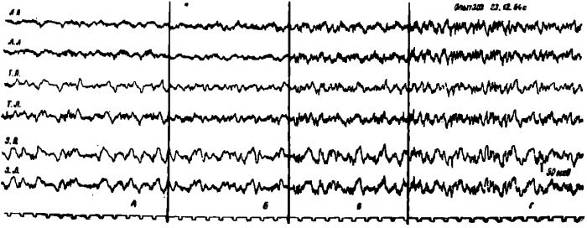
Рис. 5. Изменения электрической активности коры мозга у голодной кошки после введения ей молока в желудок. А — исходная ЭЭГ голодного состояния. Б — через 5 мин. В — через 15 мин. Г — через 40 мин. после введения молока в желудок. Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сензомоторная правая. СМЛ - сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек.
Только через час после введения пищи в желудок наблюдалось устойчивое возрастание амплитуды биотоков и устранение «голодной» активации коры мозга.
Все эти опыты указывали на то, что наблюдаемая у голодных животных под уретановым наркозом избирательная регионарная активация передних отделов коры мозга обусловливалась восходящими активирующими влияниями пищевого центра.
Последнее изменение этой страницы: 2016-06-09
lectmania.ru. Все права принадлежат авторам данных материалов. В случае нарушения авторского права напишите нам сюда...