Категории:
ДомЗдоровьеЗоологияИнформатикаИскусствоИскусствоКомпьютерыКулинарияМаркетингМатематикаМедицинаМенеджментОбразованиеПедагогикаПитомцыПрограммированиеПроизводствоПромышленностьПсихологияРазноеРелигияСоциологияСпортСтатистикаТранспортФизикаФилософияФинансыХимияХоббиЭкологияЭкономикаЭлектроника
Распространение восходящих активирующих влияний гипоталамуса на синаптические организации коры мозга у голодных животных
Приступая к настоящей серии исследований, мы решили в качестве показателя функциональной активности синаптических организаций коры мозга использовать вызванный потенциал, возникающий в коре мозга в ответ на одиночное раздражение седалищного нерва.
Предыдущими исследованиями сотрудников нашей лаборатории (3, 4, 10, 13 и др.) было показано, что вызванный потенциал, регистрируемый с поверхности коры мозга, представляет собой результат множественных восходящих влияний, поступающих к синапсам корковых клеток по различным каналам от различных подкорковых образований.
Согласно этим представлениям, отрицательная фаза и вторичные ответы вызванного потенциала обусловливаются в коре множественными восходящими влияниями неспецифических подкорковых систем, которые адресуются к аксодендритным синапсам поверхностного плексиморфного слоя. В то же время положительная фаза первичного ответа определяется возбуждениями аксосоматических сипапсов на нейронах IV слоя коры за счет прихода возбуждений по классическим лемнисковым путям (2, 3 и др.).
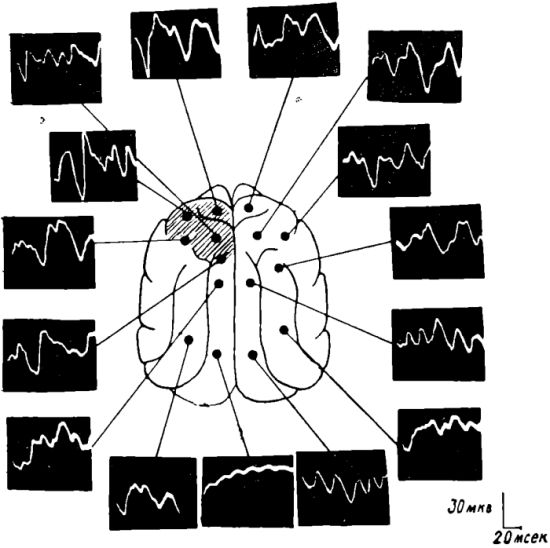

Рис. 11. ЭЭГ (внизу) и карта, показывающая отсутствие распространения соматосензорных вызванных потенциалов на кору мозга у кошки после 2-суточного голодания. Уретановый наркоз.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сензомоторная правая, СМЛ — сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗГТ — затылочная правая, ЗЛ — затылочная левая обл.
Отметка времени 1 сек.
Приступая к настоящей серии экспериментом, мы исходили из следующего предположения. Состояние голода на основе восходящих активирующих влияний подкорковых образований, создает в центральной нервной системе определенное доминантное состояние, сопровождающееся мобилизацией определенных корковых синаптических образований. В этом состоянии любое возбуждение другой биологической модальности, поступающее к коре мозга, должно было неизбежно вступить в какие-то взаимодействия с наличным пищевым возбуждением и, естественно, претерпеть определенные изменения. Особенно отчетливо эти изменения должны быть выражены у соматосензорного вызванного ответа, поскольку формирующее его возбуждение адресуется к тем же передним отделам -коры мозга, куда поступают и возбуждения пищевого центра.
Проведенные опыты показали следующее. У голодных животных, находящихся под уретановым наркозом, соматосензорные вызванные потенциалы в ответ на одиночное раздражение седалищного нерва регистрировались только в ограниченном участке сензомоторной области коры мозга, т. е. в той области, которая является специфической проекционной зоной соматосензорных возбуждений.
Как правило, в этом пункте регистрировались вызванные ответы только с первичноположительной фазой. В ряде опытов у голодных животных соматосензорные вызванные ответы вообще не регистрировались. Характерно, что в то же время и в том же пункте коры вызванные потенциалы, возникающие у голодных животных при раздражении пищевых центров гипоталамуса, выявлялись в коре мозга совершенно отчетливо.

Рис. 12. Вызванные потенциалы в сензомоторной области коры у голодных кошек, находящихся под уретановым наркозом, при раздражении седалищного нерва (А) и различных отделов гипоталамуса 1Б.


Pиc. 13. ЭЭГ (внизу) и карта распространения соматосензорного вызванного потенциала по коре мозга кошки, накормленной перед опытом. Уретановый наркоз.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая. СМП — сензомоторная правая. СМЛ — сензомоторная левая, ТП — теменная правая, TЛ — теменная левая, ЗП — затылочная правая, 3Л — затылочная левая обл. Отметка времени 1 сек.
Наоборот, у накормленных перед опытом животных соматосензорные вызванные ответы с отчетливо выраженными первичной положительной и отрицательной фазами и нередко с вторичным положительным колебанием регистрировались над обширной областью передних отделов коры мозга.
Аналогичный эффект возрастании амплитуды отрицательной фазы в фокусе максимальной активности и расширения зоны регистрации вызванных ответов наблюдался при введении голодным животным раствора глюкозы или при -коагуляции медиальных отделов гипоталамической области.

Рис. 14. Изменения характера выявления и распространения соматосензорного вызванного ответа по коре мозга у голодной кошки после внутривенного введения 40% — 2 мл раствора глюкозы. Уретановый наркоз.
I — вызванный ответ в тех же точках 1, 2, 3 до введения глюкозы; II — вызванный ответ в тех же точках через 15 минут после введения глюкозы.
Все это указывало на то, что изменения соматосензорного вызванного ответа у голодного животного определялись восходящими активирующими влияниями гипоталамуса на кору мозга.
В результате этих пищевых возбуждений происходит своеобразное ««вычитание» синапсов из целостной деятельности, участвующей в формировании соматосензорных вызванных потенциалов.
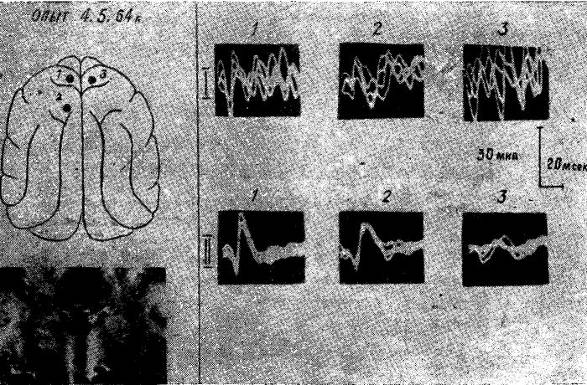
Рис. 15. Изменение характера и распространения соматосензорного вызванного ответа по коре мозга у голодной кошки после коагуляции вентромедиальных отделов гипоталамуса и субталамуса. Уретановый наркоз.
I — исходный вызванный потенциал в точках 1, 2, 3. II — вызванный потенциал в тех же точках после коагуляции медиальных отделов гипоталамуса и субталамуса.
Поскольку в состоянии голода у животных наблюдалось избирательное подавление отрицательной фазы первичного ответа и вторичного положительного колебания соматосензорного вызванного ответа, то можно думать, что возбуждения гипоталамических отделов пищевого центра в состоянии голода адресуются именно к тем синаптическим организациям сензомоторной области коры, которые формируют эти фазы вызванного потенциала. Так как отрицательная фаза первичного ответа, а также вторичные потенциалы обусловливаются восходящими влияниями неспецифических подкорковых систем, адресующихся к аксодендритньим синапсам поверхностных слоев коры (2, 3, 4 и др.), можно думать, что восходящие активирующие влияния гипоталамических отделов пищевого центра специфически адресуются именно к этим синаптическим образованиям коры головного мозга.
Специальные опыты показали, что блокада распространения самотосензорных потенциалов по коре мозга у голодных животных происходит и на уровне медиальных ядер таламуса. Коагуляция медиальных ядер таламуса у накормленных животных прекращает генерализованное распространение вызванных потенциалов на передние отделы контралатерального полушария.
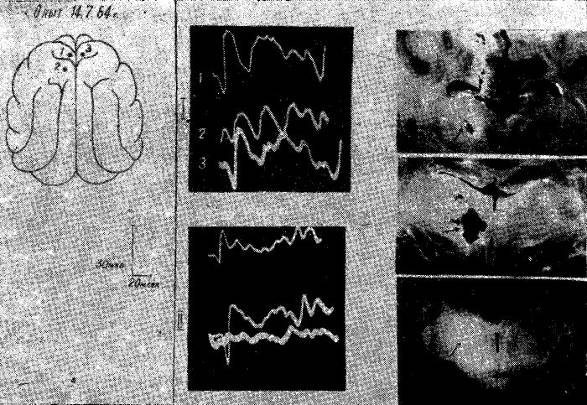
Рис. 16. Изменения распространения соматосензорного вызванного потенциала по коре мозга у накормленной кошки. Уретановый наркоз.
I — до, II — после коагуляции медиальных групп ядер правого таламуса. После этого вызванный потенциал перестает распространяться на ипсилатеральной стороне.
На основании этих опытов мы пришли к выводу, что восходящие активирующие влияния гипоталамуса в состоянии голода специфически распространяются и на аксодендритные синапсы передних отделов коры за счет первичного возбуждения медиальной группы таламических ядер.
Последнее изменение этой страницы: 2016-06-09
lectmania.ru. Все права принадлежат авторам данных материалов. В случае нарушения авторского права напишите нам сюда...