Категории:
ДомЗдоровьеЗоологияИнформатикаИскусствоИскусствоКомпьютерыКулинарияМаркетингМатематикаМедицинаМенеджментОбразованиеПедагогикаПитомцыПрограммированиеПроизводствоПромышленностьПсихологияРазноеРелигияСоциологияСпортСтатистикаТранспортФизикаФилософияФинансыХимияХоббиЭкологияЭкономикаЭлектроника
Нейрохимические механизмы восходящих активирующих влияний на кору мозга у голодных животных
Наличие у голодных животных системы пищевого возбуждения избирательно объединяющей корковые и подкорковые элементы, поставило перед нами вопрос, какие механизмы производят функциональное объединение различных нервных элементов в единую систему пищевого возбуждения, не блокируемую даже уретаном? Мы предположили, что такое функциональное объединение при пищевом возбуждении может быть основано на едином химическом механизме системы восходящих активирующих влияний гипоталамических пищевых центров на кору головного мозга.
Эксперименты, проведенные в нашей лаборатории показали, что такое адренологическое вещество, как аминазин, избирательно блокирует механизмы восходящей активности коры при болевой реакции, но оставляет при этом относительно интактной активацию коры мозга, обусловленную «голодным» возбуждением животного (15). Это позволяло думать, что пищевое возбуждение у голодных животных определялось не адренергическими механизмами мозга. Учитывая литературные данные о том, что ретикулярная формация ствола мозга использует в своих восходящих активирующих влияниях на кору мозга и холинергические механизмы (7, 8, 9, 18, 28 и др.) на предположили, что механизмы восходящих активирующих влияний на кору мозга у голодных животных строятся на основе холинореактивных элементов. Опыты подтвердили такое предположение.

Рис. 17. Изменения ЭЭГ голодной кошки после подкожного введения 0,1%—2 мл атропина (правая часть рисунка). В правой части рисунка кроме того показан эффект раздражения седалищного нерва на фоне действия атропина. Уретановый наркоз.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сензомоторная: правая. СМЛ — сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек.
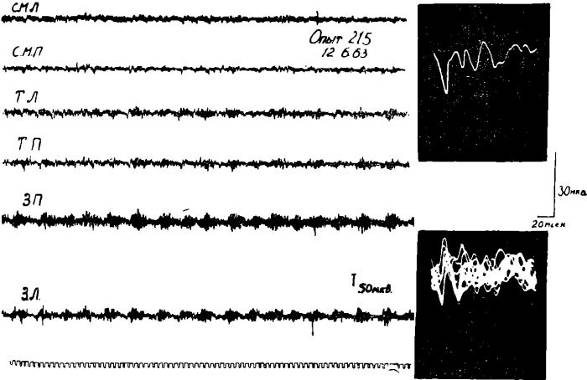

Рис. 18. Изменения характера соматосензорного вызванного потенциала, регистрируемого в сензомоторной коре I — до, II — после введения 0,1% — 2 мл атропина. Уретановый наркоз.
Так, подкожное введение голодным животным растворов атропина или амизила из расчета 0,5—0,6 мг на 1 кг веса на фоне отчетливой избирательной десинхронизации ЭЭГ в передних отделах коры мозга уже через 2—5 минут устраняло «голодное» возбуждение и приводило к появлению во всей коре мозга медленных высокоамплитудных колебаний.
Однако, пороговое раздражение седалищного нерва при этом по-прежнему вызывало, хотя и менее отчетливую, генерализованную ЭЭГ активацию всей коры мозга. После введения голодным животным холинолитических веществ у них, так же как и у накормленных животных, начинала отчетливо регистрироваться отрицательная фаза корковых соматосензорных потенциалов. Это указывало на то, что холинолитические вещества избирательно блокировали только механизмы пищевой восходящей активации коры больших полушарий, оставляя относительно интактными механизмы болевой активации. Дальнейшее подтверждение этого было получено в опытах, проведенных в нашей лаборатории (14).
Аппликация холинолитических веществ непосредственно на кору мозга у голодных животных в его опытах приводила в пункте аппликации к устранению «голодной» активации и резкому возрастанию здесь отрицательной фазы соматосензорных вызванных потенциалов. Аналогичные данные были получены и А. А. Панфиловым (1965), «который показал, что избирательная ЭЭГ активация передних отделов коры мозга у голодных животных исчезает после локальной инъекции холинолитимов непосредственно в область латерального гипоталамуса.
Все это указывало на то, что система пищевого возбуждения у животных после 1—2-суточного голодания строится преимущественно на основе холинергических механизмов.
Однако при этом следует иметь в виду, что в активации коры мозга у «голодных животных принимают участие и адренергические механизмы. Опыты показали, что адренергические механизмы вое более интенсивно возбуждаются при увеличении сроков голодания животных Наблюдаемая у животных после 4-суточного голодания генерализованная ЭЭГ активация, в отличие от региональной активации передних -отделов коры мозга у животных после 1—2-суточного голодания, блокируются аминазином. Это дает основание считать, что адренергические механизмы определяют ту реакцию общего напряжения организма, которая развивается при увеличении сроков голодания.
Заключение
Проведенные нами эксперименты показывают, что пищевое возбуждение у голодных животных охватывает целую систему функционально объединенных нервных элементов, находящихся как в подкорковых образованиях, так и в кюре головного мозга. Эта система «голодного» возбуждения формируется на основе специфических механизмов восходящих активирующих влияний подкорковых образований на кору головного мозга. Возбуждения подкорковых образований строго избирательно мобилизуют элементы коры головоного мозга, что выражается в специфической форме активации ЭЭГ голодных животных.
В механизме этих восходящих активирующих влияний на кору мозга центральная роль принадлежит пищевым центрам гипоталамичеовой области. Искусственное снижение функциональной активности пищевых центров гипоталамуса приводит к устранению пищевого возбуждения у голодных животных, как в подкорковых образованиях, так и в коре головного мозга.
Из всего этого следует, что гипоталамические отделы пищевого центра обладают огромной силой тонического воздействия на другие структуры мозга и прежде всего на кору больших полушарий. Все это дает нам основание рассматривать пищевые центры гипоталамуса как своеобразный «пейцмекер» всей системы пищевого возбуждения у голодных животных.
Проведенные опыты показывают, что распространение восходящих активирующих влияний гипоталамуса на кору мозга у голодных животных осуществляется через медиальные ядра таламуса. Наряду с этим гипоталамические пищевые центры оказывают активирующее действие на ретикулярную фармацию среднего мозга. Эти влияния все более усиливаются при увеличении сроков голодания. Вовлечение в систему пищевого возбуждения ретикулярной формации среднею мозга приводит к появлению генерализованных активирующих восходящих влияний на всю кору головного мозга, что, в свою очередь, определяет еще более высокий энергетический тонус пищедобывательного поведения.
Разобранные нами специальные механизмы восходящих активирующих влияний на кору мозга у голодных животных подтверждают представления нашей лаборатории о такой функциональной организации взаимоотношений коры и подкорки, при которой подкорковые образования берут на себя роль своеобразного источника энергии, который избирательно поддерживает в рабочем тонусе всю обширную функциональную систему данной специфичности. Очевидно, именно, через этот физиологический механизм реализует себя та подкорковая «слепая сила», о которой много раз писал И. П. Павлов.
Наши опыты показали, что восходящие активирующие влияния подкорковых областей на кору мозга при голоде за счет первичного возбуждения медиальных ядер таламуса распространяется преимущественно на аксодендритные синапсы плексиморфного слоя коры мозга. Восходящие активирующие влияния подкорковых отделав пищевого центра у голодного животного производят избирательную мобилизацию корковых синапсов из их общей массы. Только на основе избирательного возбуждения клеток коры мозга может возникнуть специальная пищедобывательная деятельность. Такое избирательное функциональное вовлечение инициативными подкорковыми пищевыми центрами в свою деятельность элементов коры мозга у голодных животных происходит на основе их единых, преимущественно холинергичеcких, механизмов.
Из всего изложенного выше следует, что пищевое возбуждение формируется на основе такой вертикальной построенной функциональной организации, в которой инициативным пунктом являются пищевые центры гипоталамуса, избирательно мобилизующие для пищедобывательной деятельности другие подкорковые аппараты, а также клетки коры головного мозга.

Рис. 19. Схема восходящих активирующих влиянии гипоталамических отделов пищевого центра на кору мозга. I — состояние голода, II — после приема пищи. Обозначения: Г — гипоталамус, Т — таламус, РФ — ретикулярная формация.
Благодаря таким вертикально организованным системам восходящих активирующих влияний подкорковых образований на кору мозга, весьма малое количество начальной энергии, которое образуется при возбуждении центральных рецепторов через мобилизацию многочисленных корковых нейронов, может привести в последующем к появлению огромной энергии. Именно в этом заключается смысл формирования этих реакций по доминантному типу.
Выводы
1. Нейрофизиологическую основу состояния голода и пищевой мотивации, определяющей пищедобывательное поведение, составляют специфические механизмы восходящих активирующих влияний подкорковых отделов пищевого центра на кору головного мозга.
Пищевое возбуждение у животных после 1—2-суточного голодания проявляется в форме избирательной активации ЭЭГ передних отделов коры мозга даже под уретановым наркозом, несмотря на глубокий сон животного.
2. Избирательная активация коры головного мозга у голодных животных определяется двойными влияниями со стороны «голодной» крови и нервных импульсаций, поступающих от желудочно-кишечного тракта по блуждающим нервам и чувствительным волокнам, идущим через спинной мозг.
3. Активация коры головного мозга у голодных животных определяется целой системой возбужденных подкорковых образований, находящихся на различных уровнях ствола мозга. Она объединяет пищевые центры гипоталамуса (латеральные и медиальные его отделы), медиальные отделы таламуса и ретикулярной формации среднего мозга. Во всех этих отделах у голодных животных под уретановым наркозом, так же как и в передних отделах коры мозга, регистрируются высокочастотная низкоамплитудная электрическая активность, характеризующая состояние возбуждения.
4. Центральным и инициативным пунктом системы возбужденных подкорковых образований, оказывающих у голодных животных восходящие активирующие влияния на кору мозга, являются гипоталамические отделы пищевого Центра. Их можно рассматривать как своеобразный «пейцмекер» всей системы пищевого возбуждения.
5. Восходящие активирующие влияния гипоталамуса на кору мозга у голодных животных распространяются преимущественно через медиальные отделы таламуса.
Наряду с восходящими активирующими влияниями на кору мозга гипоталамические отделы пищевого центра у -голодных животных оказывают активирующие влияния и на ретикулярную формацию среднего мозга. Эти влияния гипоталамических центров на ретикулярную формацию среднего мозга значительно усиливаются при увеличении сроков голодания.
6. Восходящие активирующие влияния -гипоталамуса на кору мозга в состоянии голода через первичное возбуждение медиальных ядер таламуса распространяются преимущественно на аксодендритные синапсы поверхностного плексиморфного слоя передних отделов коры мозга.
7. Функциональное объединение различных нервных элементов, находящихся в подкорковых образованиях и в коре мозга, в систему пищевого возбуждения у животных после 1—2-суточного голодания происходит на основе единых химических, преимущественно холинергических механизмов.
8. Состояние насыщения характеризуется значительным снижением функциональной активности различных отделов пищевого центра, находящихся на разных уровнях центральной нервной системы и устранением их восходящих активирующих влияний на кору мозга.
ЛИТЕРАТУРА
1. Агафонов В. Г. Журн. невропатологии и психиатрии, 1956, 56, 94.
2. Анохин П. К. Журн. высшей нервной деятельности, 1962, 12, З, 379.
3. Ата-Мурадова Ф. А. Материалы 1-ой научной конф., посвященной проблемам физиологии, морфологии и клиники ретикулярной формации головного мозга, М., 1960, стр. 14.
4. Ата-Мурадова Ф. А. Физиологический журнал СССР, 1963, 49, 7, 781.
5. Гавличек В. Физиологический журнал СССР, 1958, 44, 4, 305.
6. Зачиняева И. А. Материалы 3-ей конф. по вопросам электрофизиологии цнс, Киев, 1960, стр. 161.
7. Ильюченек Р. Ю. Сб. «Электрофизиология н. е.», Ростов-на-Дону, 1963, стр. 169.
8. Каграманов К. М. Тр. ин-та нормальной и патоуюгич. физиологии АМН СССР, 1964, т. 7, стр. 47.
9. Калюжный Л. В. Жури, высшей нервной деятельности, 1962, 12, 2, 313.
10. Лю-Джан-Гуй. Электрофизиологический анализ механизмов генерализации возбуждений в коре больших полушарий головного мозга. Канд. дисс., М., 1960.
11. Макаров Ю. А. Физиологические механизмы взаимодействия условных реакций различного биологического качества. Канд. дисс., М., 1960.
12. Панфилов А. А. и Лосева Т. Н. Материалы 26 итоговой сессии научн. студенч. общества 1 МОЛМИ, М., 1964, стр. 18—20.
13. Полянцев В. А. и Сербиненко М. В. Сборник «Вопросы физиологии и патологии н. е.». Тр. института нормальной и патологической физиологии АМН СССР, М., 1962, т. 6, стр. 8—10.
14. Туренко А. И. Сб. «Физиология и патология н. е.». Тр. института нормальной и патологической физиологии АМН СССР, 1964, т. 7, стр. 100.
15. Фадеев Ю. А, Рефераты работ Всесоюзной конф. молодых ученых ин-та нормальной и патологической физиологии АМН СССР, М., 1956, 56, 116.
16. Шумилина А. И. Журн. невропатологии и психиатрии им. Корсакова, 1956, 56, 116.
17. Anand В. К. and Brobeck J. R. Vale J. Biol Med., 1951, 24 123
18. Bradley P. B. and Eccles G. С .J. Physiol. (Zond), 1953, 120, 14 p.
19. Brobeck J. K- Am. N. G. Acad. Sci., 1955, 63, 44.
20. J a s p e r H. H. and A j m о n e—M a r s a n C. Nat. Research. Council, of Canada Ottawa, 1952.
21. J a s p e r H. H. Neurophysiol., 1949, 1, 405.
22. Larsson S. Acta Physiol. Scand., 1954, 32, suppi 115.
23. M a g e r J. J. Bull. New Engl. Med. Cent., 1952, 14, 43.
24. Moruzzi G. und Magoun H. W. Neurophysio)., 1949, 1, 455.
25. M u r p h у J. P. and G e 1 1 h о r n E. J. Neurophysiol., 1945, S, 341.
26. Q u z m a n F. С., A l с a r a z M. and Fernandez-Guar-d i о 1 a A. Boletin inst. Egfud. Med Biol., 1858, 16, 1, 29.
27. Rothballer A. B. Neurophys., 1956, 8, 603.
28. R i n a 1 d i F. and Hi m w i с h H. E. Arch. Neurol. Psychiat., 1955, 73, 396.
Последнее изменение этой страницы: 2016-06-09
lectmania.ru. Все права принадлежат авторам данных материалов. В случае нарушения авторского права напишите нам сюда...