Категории:
ДомЗдоровьеЗоологияИнформатикаИскусствоИскусствоКомпьютерыКулинарияМаркетингМатематикаМедицинаМенеджментОбразованиеПедагогикаПитомцыПрограммированиеПроизводствоПромышленностьПсихологияРазноеРелигияСоциологияСпортСтатистикаТранспортФизикаФилософияФинансыХимияХоббиЭкологияЭкономикаЭлектроника
Механизмы регуляции поведения у собак при наличии у них голода и жажды
Н. Я. АСМАЯН (Москва)
При изучении высшей нервной деятельности за последнее время все больше внимания уделяется механизмам регуляции свободного поведения животных.
Исследования в этом направлении в Советском Союзе уже давно проводились во многих лабораториях (1, 4).
Для изучения поведения у собак были предложены различные методики, но среди них предпочтение следует отдать методике П. К. Анохина, поскольку она в отличие от всех остальных полностью сохраняет все принципы условно-секретной методики И. П. Павлова и является ее дальнейшим усовершенствованием. По этой методике выработка условного рефлекса проводится также в звуконепроницаемой камере, где, однако, животное может свободно перемещаться по станку и получать подкрепление условного раздражителя на двух его сторонах. Помимо слюноотделения по этой, методике регистрируются движения и дыхание животного.
Использование этого метода уже с самого начала позволило выявить некоторые особенности корковой деятельности.
Прежде всего было показано, что условнорефлекторная деятельность есть целостная реакция всего организма, в которой деятельность отдельных органов и систем не однородна. Если деятельность одних отражает специфическую активность, направленную на действие раздражителя, например, слюноотделение при приеме пищи, то деятельность других — общую активность, направленную на подготовку условий для удовлетворения потребности организма, например, подготовки условий для приема пищи, изменение дыхания, уровня кровяного давления и сердечной деятельности и, наконец, двигательная реакция, подход к кормушке и т. д.
На основании этих фактов возникло представление о функциональной системе, как целостной реакции организма на раздражение, направленной на достижение различных приспособительных эффектов организма (П. К. Анохин, 1935). Каждая функциональная система имеет характерные узловые пункты: афферентный синтез, акцептор действия, обратная афферентация от результатов действия и т. д.
Известно, что афферентный синтез возникает в результате анализа всей поступающей в центральную нервную систему информации. Информация может поступать как извне, в виде пусковой и обстановочной афферентации, так и изнутри, как информация об изменении констант или увеличения той или другой потребности организма.
Появление потребности вызывает стремление к ее ликвидации. Такое стремление к удовлетворению основных потребностей организма (голод, жажда и т, д,) получили название мотивации или основных влечений организма.
В результате афферентного синтеза выбор ценной информации всегда-происходит строго в соответствии с потребностью организма и возникшей на ее основе мотивации. Поэтому мотивация или влечение являются главной движущей силой, которая хотя и формируется на основе афферентного синтеза, оказывает основное влияние на образование функциональной системы и на поведение животного.
Подтверждением этого может служить тот факт, что выработка пищевых условных рефлексов возможна лишь у голодных собак, когда достаточно выражена пищевая мотивация. В случае, если пищевая мотивация отсутствует или ослаблена, возможна другая доминантная деятельность, другой процесс в афферентном синтезе и образование другой функциональной системы.
В настоящее время хорошо изучены механизмы образования пищевой функциональной системы. Изучена роль в ее образовании пусковой и обстановочной афферентации. Однако значение мотивации, как основной движущей силы в процессе формирования функциональной системы и особенно поведения, изучено недостаточно. Известно лишь, что по мере удовлетворения потребности организма, мотивация уменьшается, а в связи с этим происходят изменения в афферентном синтезе, в функциональной системе и, наконец, в самом поведении животного.
Отсутствие достаточных сведений о роли мотиваций в формировании поведения животного зависит, по-видимому, от того, что при его изучении для животного чаще всего создавались условия, когда усиливалась только одна потребность и, следовательно, была выражена только одна мотивация. От всех остальных потребностей животное ограждалось и если они обнаруживались, то по возможности удовлетворялись без особого учета. Так, например, известно, что при работе в камере условных рефлексов животные специально получали воду с тем, чтобы жажда не оказывала влияния на их пищевое поведение.
Такая форма исследования, когда все внимание сосредоточено на формировании только одной функциональной системы имело большое значение при изучении механизмов образования пищевых условных рефлексов, однако, она недостаточна и ограничивает возможность анализа тех отношений, которые могут возникнуть в естественных условиях, когда постоянно возникает необходимость в удовлетворении не одной, а нескольких потребностей, когда одна мотивация может сменить другую, а поведение животного неожиданно и резко изменяется.
Таким образом, чтобы понять механизмы регуляции поведения, необходимо в эксперименте создать такие условия, при которых у животного могла быть не одна потребность и не одна мотивация.
Исходя из этого, в настоящей работе была поставлена задача провести исследование механизмов регуляции поведения на собаках, при условии наличия у них одновременно двух мотиваций: мотивации голода и мотивации жажды.
Для проведения опытов была использована методика выработки условных рефлексов в камере конструкции П. К. Анохина с двухсторонним подкреплением. Удовлетворение мотивации голода и жажды строго фиксировалось за кормушками. В кормушке на левой стороне всегда подавалась вода (40 см3), на правой стороне — сухари (20 г). Сигнал, предупреждающий о возможности получения подкрепления для обеих кормушек был один и тот же — метроном — 60 ударов в минуту.
Вначале у животных вырабатывался обычный условный рефлекс на М-60 при подаче обеих кормушек, так что животное устремлялось к одной из них по собственному выбору.
Эти условные рефлексы были выработаны сравнительно легко. Обе собаки стали бегать к той и другой кормушке на сигнал уже спустя 10—12 дней.
Для поддержания у собак весом в 16—17 кг умеренного голода и умеренной жажды был установлен пищевой рацион на сутки: 150 г ржаного хлеба, 150 г сырого мяса, 75 г белых сухарей и 100 мл воды. Опыты ставились ежедневно, спустя 18—20 часов после приема пищи. Для поддержания у собак умеренного голода и сильной жажды был использован тот же пищевой рацион, но без воды. Опыты также начинались спустя 18—20 часов после приема пищи, но проводились через день. Воду животные получали только во время опыта. Причем им предоставлялась возможность пить до отказа. Собаки, находясь на этих рационах, были в хорошем состоянии и в весе не теряли.
После того, как указанные условные рефлексы были выработаны, мы в схему их выработки ввели некоторое инструментальное дополнение. Оно заключалось в том, что та и другая кормушки на сигнал экспериментаторами не поддавались, а должны были запускаться самой собакой в результате нажатия ею по собственному выбору на соответствующий для каждой кормушки рычаг. Оба рычага имели определенные внешние различия: для «водяной» кормушки площадка рычага имела форму креста, а для «сухарной» — форму круга. На станке оба рычага могли свободно переставляться.
В целях объективного контроля за поведением животного велась регистрация его реакций, подаваемых сигналов и подкрепления.
Все это регистрировалось на бумажной ленте с помощью чернильных самописцев лентопротяжного механизма. Для регистрации подхода к кормушкам, еды сухарей и питья воды сверху кормушек были пристроены, специальные коробки, которые прикрывали отверстия кормушек. Передняя стенка коробки висела на петле, так что животное могло ее легко отодвигать мордой и проникать к воде или сухарям. Движение передней стенки коробки использовалось для регистрации момента обследования кормушки, еды сухарей и питья воды собакой Отодвигание передней стенки коробки на «сухарной» кормушке регистрировалось на бумаге, как подъем писчика вверх, а отодвигание передней стенки коробки «водяной» кормушки, как опускание этого же писчика вниз.
Движение животного по станку регистрировалось при помощи другого писчика. Расположение животного на середине станка писалось как нулевая линия. Движение в сторону сухарной кормушки отмечалось движением писчика вверх, а движение в сторону водяной кормушки — как движение писчика вниз. Кроме этого, регистрировались отметки подачи условного сигнала, отметка момента и сила нажатия собакой на рычаг и отметка подачи кормушки. Нажатие собакой на сухарный рычаг регистрировалось подъемом писчика вверх, нажатие на водный рычаг — опускание вниз. Кроме того, обычным капельно-контактным методом велась регистрация слюноотделения и регистрировалось дыхание при помощи пневматической передачи.
Сначала, когда собаки приучались к рычагам, последние устанавливались на станке, а в дальнейшем ставились в определенных местах на приставном столике.
Опыты показал, что животные довольно быстро научились давить на площадку рычага, в результате чего запускали кормушку и получали соответствующее подкрепление.
При этом следует отметить, что как в условиях умеренной жажды и умеренного голода, так и сильной жажды и умеренного голода, животные оказываются сосредоточенными на удовлетворении главным образом одной мотивации — мотивации голода или мотивации жажды. В соответствии с этим они определяют нужный рычаг и пользуются им в дальнейшем.
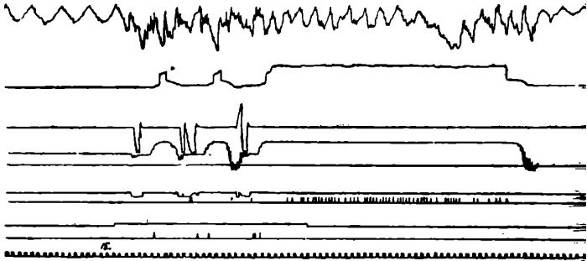
Рис. 1. Поиск нужного рычага для удовлетворения мотивации голода.
Обозначения сверху вниз: а — кривая дыхания; б — отметка обследования собакой кормушки и еда сухарей; в — отметка нажатия собакой на рычаг — вниз на «водяной», вверх на «сухарный»; г — отметка движения собаки по станку, от средней линии вверх — в сторону сухарной кормушки, вниз — в сторону водяной; д — отметка перехода на приставной столик; е — регистрация слюноотделения в каплях; ж — отметка подачи условного сигнала (М-60 в 1 мин); з — отметка подачи кормушки; и — отметка времени в сек.
В условиях умеренного голода и умеренной жажды животные больше ориентируются на кормушку с пищевым подкреплением и своим поведением характеризуют стремление к удовлетворению пищевой мотивации. На сигнал они бегут либо прямо к «сухарному» рычагу, либо иногда прежде всего к «сухарной» кормушке и уже после ее обследования к рычагу. Здесь они нажимают на него, запускают кормушку, вновь бегут к ней и поедают пищу. Иногда животное может ошибиться и нажать не тот рычаг, который вызывает погребное для него подкрепление, тогда, несмотря на звук от срабатываемой другой кормушки, оно все же направляется к той стороне, где ожидается пищевое подкрепление. Проверка кормушки подтверждает ошибку и стимулирует поиск.
В случае умеренного голода и резко выраженной жажды животные, наоборот, больше сосредоточивают свое внимание на «водяной» кормушке. Они постоянно находятся на ее стороне, редко забегают на сторону «сухарной» кормушки, а в случае ее подачи и подхода к ней отказываются от еды.
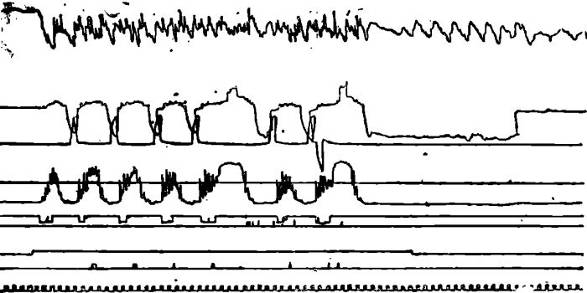
Pис. 2. Поиск нужного рычага для удовлетворения мотивации жажды.
Обозначения те же, что на рис. 1.
По мере удовлетворения этой резко выраженной мотивации жажды выявляется и все более усиливается мотивация голода. Животные все чаще начинают забегать на сторону «сухарной» кормушки. Вначале только после подкрепления мотивации жажды, затем — до и после. К этому времени они во время пауз все больше находятся на стороне «сухарной» кормушки и начинают поиск нужного рычага именно с этого места. Наконец, когда мотивация жажды оказывается удовлетворенной полностью, животное свое внимание целиком переключает на «сухарную» кормушку.
Наблюдая за поведением животного в условиях переключения с одной мотивации на другую, мы обнаружили в нем не только автоматизированные движения, но и ориентировочно-исследовательскую реакцию, связанную с сопоставлением полученных результатов с ожидаемыми.
В качестве примера приводятся данные, полученные на собаке Дружок.

Pис. 3. Исправление собакой ошибки нажатия на «водяной» рычаг при отсутствии жажды под влиянием «акцептора действия», обеспечивающего предвидение результатов, связанных со стуком водяной кормушки. Обозначения те же.
По ходу опыта собака в результате многократного подкрепления мотивации жажды переключилась на удовлетворение мотивации голода. На поданный условный раздражитель она бежит на приставной столик, но в силу автоматизированной реакции и плохой дифференциации рычагов нажимает, как и прежде, на водяной рычаг. Нажав на рычаг, собака перебегает на станок, в сторону сухарной кормушки, но в это время срабатывает водяная кормушка. На стук водяной кормушки животное останавливается, поворачивается, но к водяной кормушке не идет, а вновь перебегает на приставной столик и нажимает уже на сухарный рычаг, после этого вновь бежит в сторону сухарной кормушки. «Ожидаемый» стук сухарной кормушки совпадает с действительной. Собака подбегает к этой кормушке и получает подкормку.
Наблюдения показывают, что поведение животного в этих условиях протекает не как цепной рефлекс, а значительно сложнее. Оно оказывается зависимым не только от условных раздражителей, пусковой и обстановочной афферентации, но и от взаимодействия между потребностями и смены одной мотивации на другую.
В этом опыте обнаруживается, что со сменой мотивации изменяется ориентировка животного, изменяется и «ожидание» будущих результатов действия. Теперь животное, нажимая на рычаг, ориентируется на стук уже не «водяной», а «сухарной» кормушки.
Рассогласование между действительным стуком «водяной» кормушки и «ожидаемым» стуком «сухарной» кормушки оказывает влияние на его поведение. Животное оценивает это несоответствие между сигналом и подкреплением, вновь возвращается к рычагам и исправляет «ошибку».
Уже на основании этих фактов можно сделать вывод, что в основе регуляции поведения животного лежат более сложные механизмы мозга, чем обычная условнорефлекторная деятельность. Поведение, как видно, зависит не только от мотиваций, действия условного раздражителя и обстановочной афферентации, но и от возникающего в мозгу собаки представления о будущих результатах действия (акцептор действия).
ЛИТЕРАТУРА
1. Анохин П. К. Внутреннее торможение как проблема физиологии, Медгиз—М., 1958.
2. Беритов И. С. Нервные механизмы поведения высших позвоночных животных, М., 1961.
3. Купалов П. С., Воеводина О. Н., Волкова В. Д., Милюкова И. В., Селиванова А. Т., Сыренский В. И., Хананашвили М. М., Шичко Г. А. Ситуационные условные рефлексы у собак в норме и патологии, Изд. «Медицина», Л., 1964.
4. Протопспов В. П. Исследование высшей нервной деятельности в естественном эксперименте, Киев, 1950.
Последнее изменение этой страницы: 2016-06-09
lectmania.ru. Все права принадлежат авторам данных материалов. В случае нарушения авторского права напишите нам сюда...